

SRm160 splicing coactivator promotes transcript 3'-end cleavage
- PMID: 11739730
- PMCID: PMC134228
- DOI: 10.1128/MCB.22.1.148-160.2002
SRm160 splicing coactivator promotes transcript 3'-end cleavage
Abstract
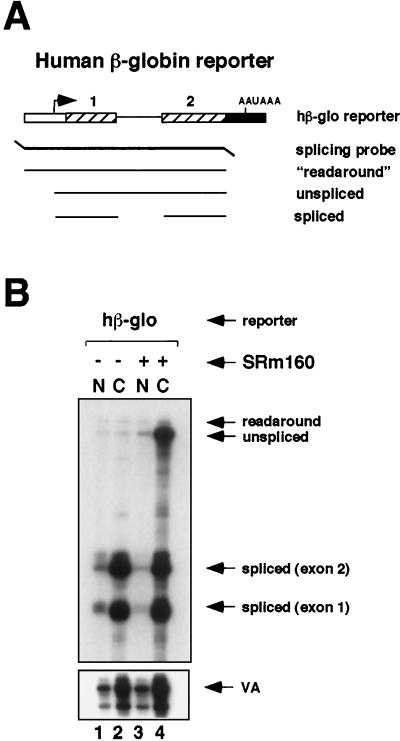
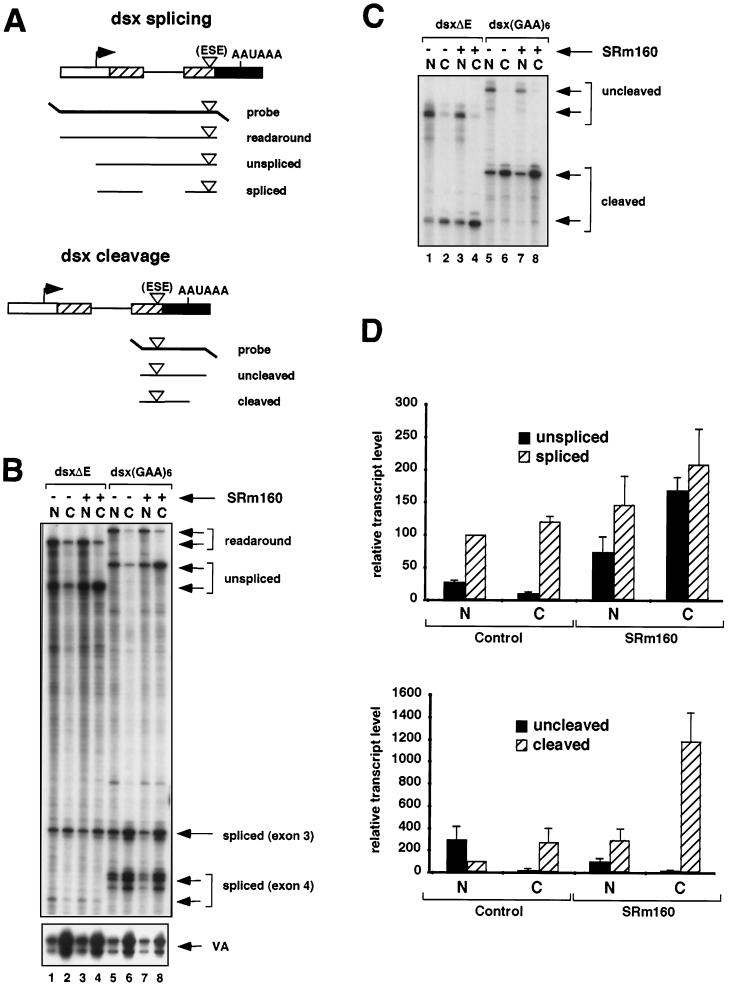
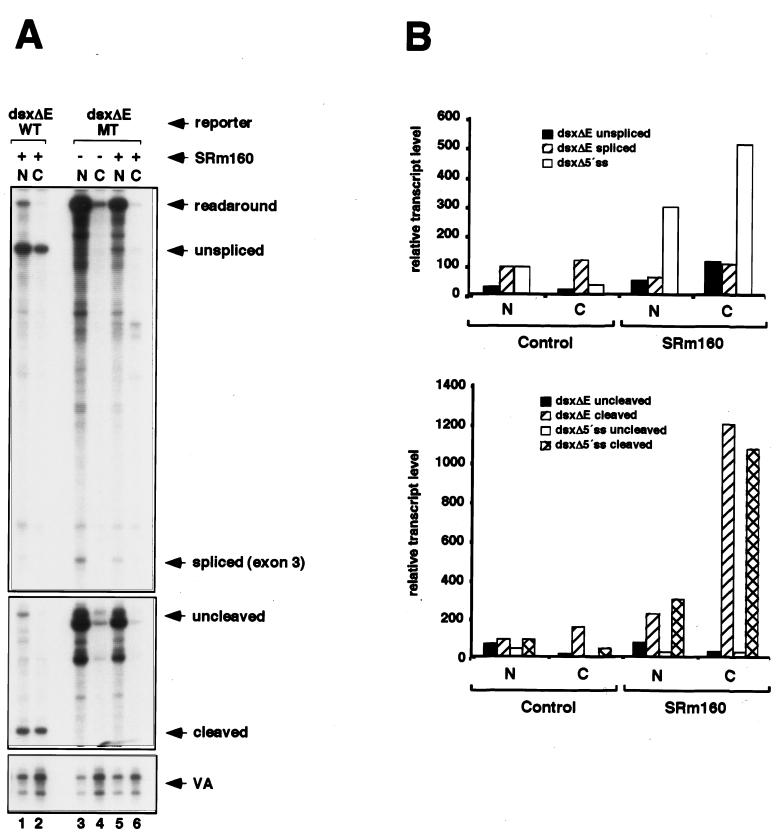
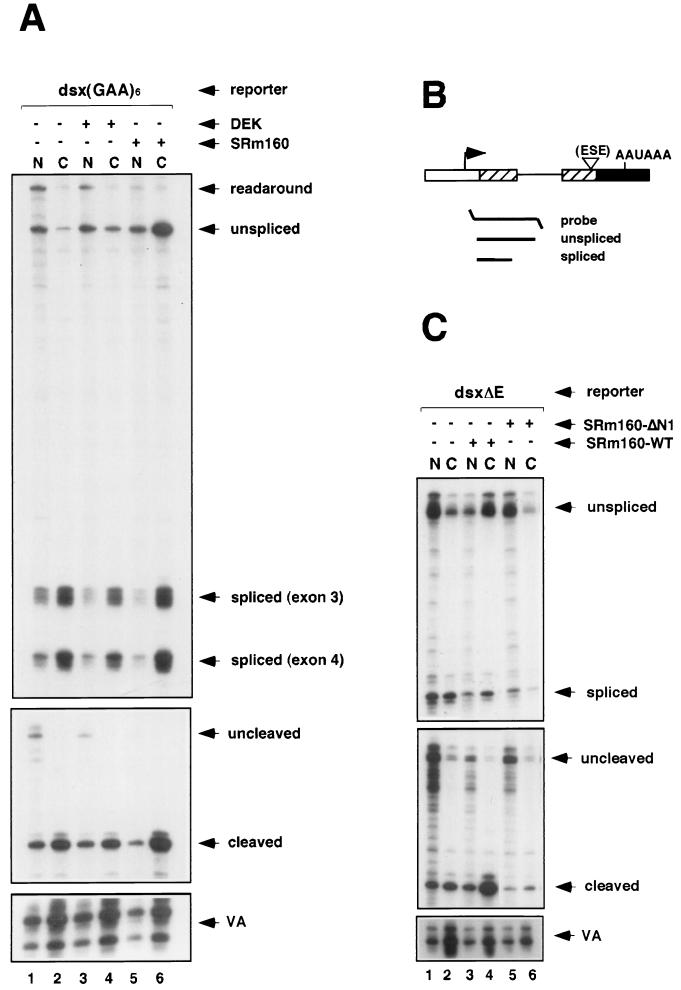
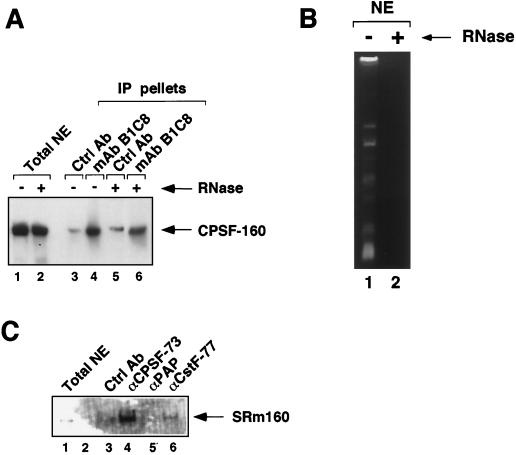
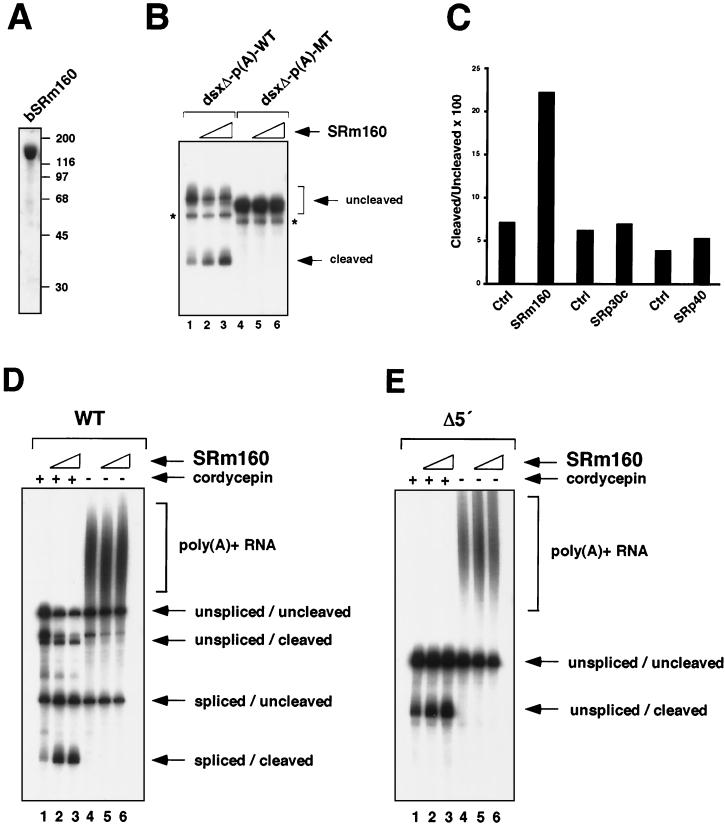
Individual steps in the processing of pre-mRNA, including 5'-end cap formation, splicing, and 3'-end processing (cleavage and polyadenylation) are highly integrated and can influence one another. In addition, prior splicing can influence downstream steps in gene expression, including export of mRNA from the nucleus. However, the factors and mechanisms coordinating these steps in the maturation of pre-mRNA transcripts are not well understood. In the present study we demonstrate that SRm160 (for serine/arginine repeat-related nuclear matrix protein of 160 kDa), a coactivator of constitutive and exon enhancer-dependent splicing, participates in 3'-end formation. Increased levels of SRm160 promoted the 3'-end cleavage of transcripts both in vivo and in vitro. Remarkably, at high levels in vivo SRm160 activated the 3'-end cleavage and cytoplasmic accumulation of unspliced pre-mRNAs, thereby uncoupling the requirement for splicing to promote the 3'-end formation and nuclear release of these transcripts. Consistent with a role in 3'-end formation coupled to splicing, SRm160 was found to associate specifically with the cleavage polyadenylation specificity factor and to stimulate the 3'-end cleavage of splicing-active pre-mRNAs more efficiently than that of splicing-inactive pre-mRNAs in vitro. The results provide evidence for a role for SRm160 in mRNA 3'-end formation and suggest that the level of this splicing coactivator is important for the proper coordination of pre-mRNA processing events.
Figures
References
-
- Birse, C. E., L. Minvielle-Sebastia, B. A. Lee, W. Keller, and N. J. Proudfoot. 1998. Coupling termination of transcription to messenger RNA maturation in yeast. Science 280:298–301. - PubMed
-
- Blencowe, B. J., A. L. Bowman, S. McCracken, and E. Rosonina. 1999. SR-related proteins and the processing of messenger RNA precursors. Biochem. Cell Biol. 77:277–291. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous