

Distinct domains of erythroid Krüppel-like factor modulate chromatin remodeling and transactivation at the endogenous beta-globin gene promoter
- PMID: 11739731
- PMCID: PMC134232
- DOI: 10.1128/MCB.22.1.161-170.2002
Distinct domains of erythroid Krüppel-like factor modulate chromatin remodeling and transactivation at the endogenous beta-globin gene promoter
Abstract
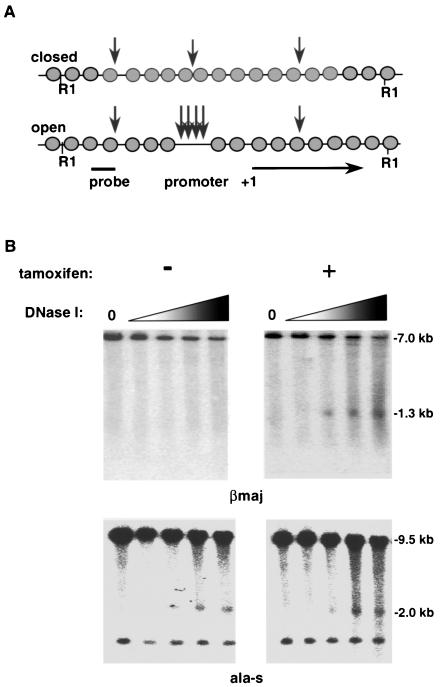
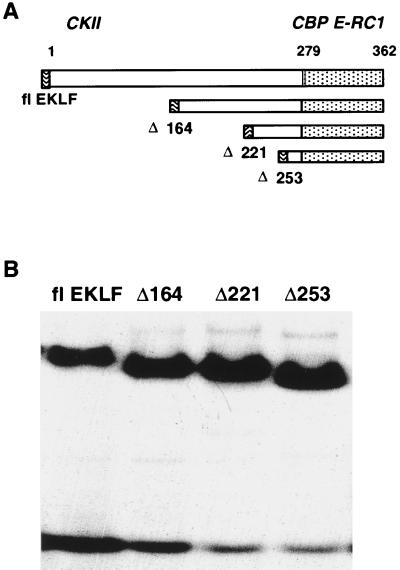
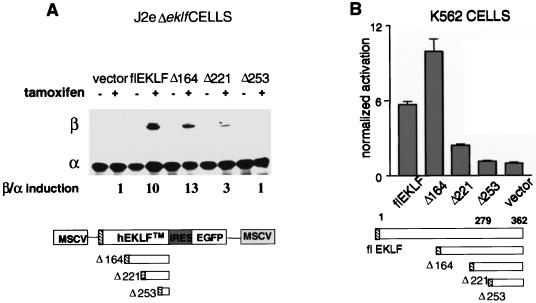
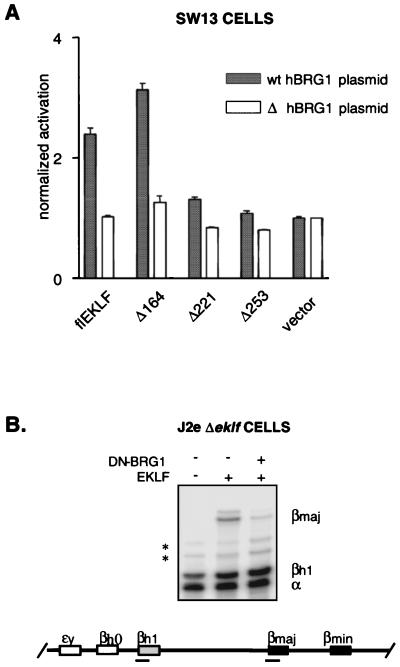
Characterization of the mechanism(s) of action of trans-acting factors in higher eukaryotes requires the establishment of cellular models that test their function at endogenous target gene regulatory elements. Erythroid Krüppel-like factor (EKLF) is essential for beta-globin gene transcription. To elucidate the in vivo determinants leading to transcription of the adult beta-globin gene, functional domains of EKLF were examined in the context of chromatin remodeling and transcriptional activation at the endogenous locus. Human EKLF (hEKLF) sequences, linked to an estrogen-responsive domain, were studied with an erythroblast cell line lacking endogenous EKLF expression (J2eDeltaeklf). J2eDeltaeklf cells transduced with hEKLF demonstrated a dose-dependent rescue of beta-globin transcription in the presence of inducing ligand. Further analysis using a series of amino-terminal truncation mutants of hEKLF identified a distinct internal domain, which is sufficient for transactivation. Interestingly, studies of the chromatin structure of the beta-promoter revealed that a smaller carboxy-terminal domain generated an open promoter configuration. In vitro and in vivo binding studies demonstrated that this region interacted with BRG1, a component of the SWI/SNF chromatin remodeling complex. However, further study revealed that BRG1 interacted with an even smaller domain of EKLF, suggesting that additional protein interactions are required for chromatin remodeling at the endogenous beta-promoter. Taken together, our findings support a stepwise process of chromatin remodeling and coactivator recruitment to the beta-globin promoter in vivo. The J2eDeltaeklf inducible hEKLF system will be a valuable tool for further characterizing the temporal series of events required for endogenous beta-globin gene transcription.
Figures



References
-
- Ansari, A. Z., J. E. Bradner, and T. V. O’Halloran. 1995. DNA-bend modulation in a repressor-to-activator switching mechanism. Nature 374:371–375. - PubMed
-
- Armstrong, J. A., J. J. Bieker, and B. M. Emerson. 1998. A SWI/SNF-related chromatin remodeling complex, E-RC1, is required for tissue-specific transcriptional regulation by EKLF in vitro. Cell 95:93–104. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous