

Repair of chromosome ends after telomere loss in Saccharomyces
- PMID: 11739802
- PMCID: PMC60777
- DOI: 10.1091/mbc.12.12.4078
Repair of chromosome ends after telomere loss in Saccharomyces
Abstract
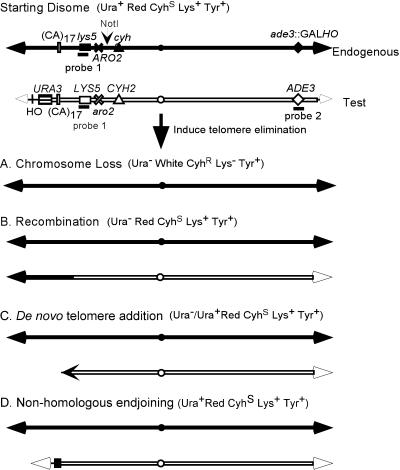
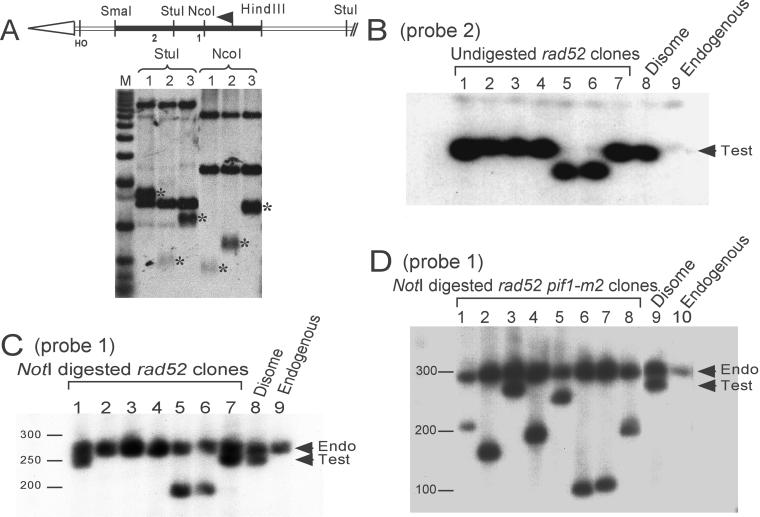
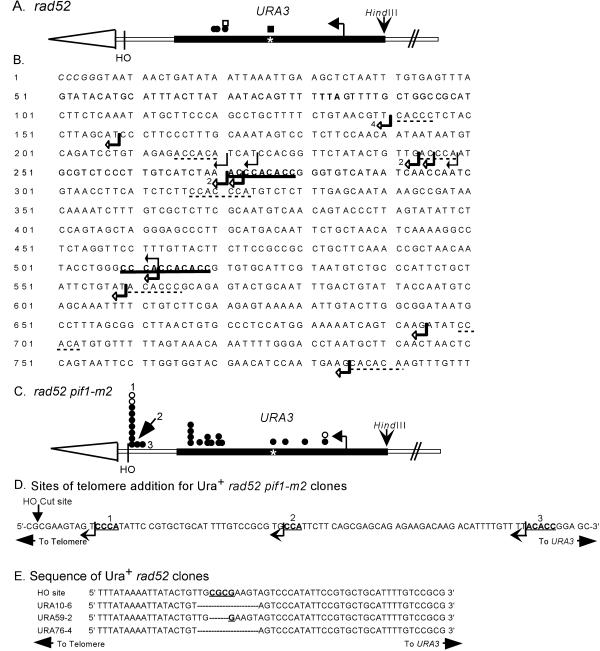
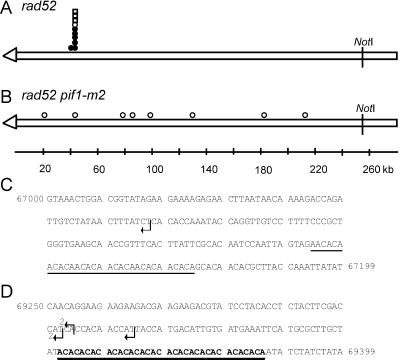
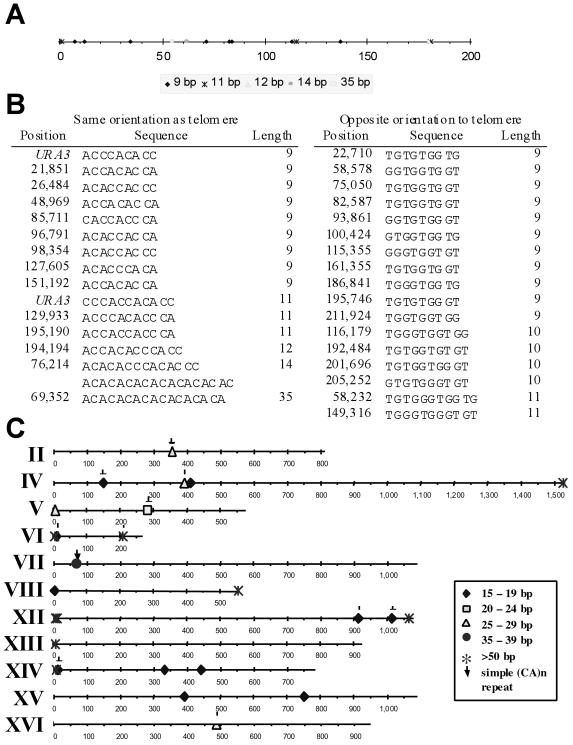
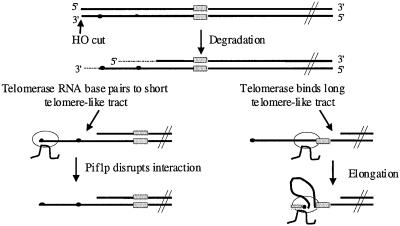
Removal of a telomere from yeast chromosome VII in a strain having two copies of this chromosome often results in its loss. Here we show that there are three pathways that can stabilize this broken chromosome: homologous recombination, nonhomologous end joining, and de novo telomere addition. Both in a wild-type and a recombination deficient rad52 strain, most stabilization events were due to homologous recombination, whereas nonhomologous end joining was exceptionally rare. De novo telomere addition was relatively rare, stabilizing <0.1% of broken chromosomes. Telomere addition took place at a very limited number of sites on chromosome VII, most occurring close to a 35-base pair stretch of telomere-like DNA that is normally approximately 50 kb from the left telomere of chromosome VII. In the absence of the Pif1p DNA helicase, telomere addition events were much more frequent and were not concentrated near the 35-base pair tract of telomere-like DNA. We propose that internal tracts of telomere-like sequence recruit telomerase by binding its anchor site and that Pif1p inhibits telomerase by dissociating DNA primer-telomerase RNA interactions. These data also show that telomeric DNA is essential for the stable maintenance of linear chromosomes in yeast.
Figures
Similar articles
-
Ku interacts with telomerase RNA to promote telomere addition at native and broken chromosome ends.Genes Dev. 2003 Oct 1;17(19):2384-95. doi: 10.1101/gad.1125903. Epub 2003 Sep 15. Genes Dev. 2003. PMID: 12975323 Free PMC article.
-
Pif1p helicase, a catalytic inhibitor of telomerase in yeast.Science. 2000 Aug 4;289(5480):771-4. doi: 10.1126/science.289.5480.771. Science. 2000. PMID: 10926538
-
Chromosome healing by de novo telomere addition in Saccharomyces cerevisiae.Mol Microbiol. 2006 Mar;59(5):1357-68. doi: 10.1111/j.1365-2958.2006.05026.x. Mol Microbiol. 2006. PMID: 16468981 Review.
-
The yeast Pif1p helicase removes telomerase from telomeric DNA.Nature. 2005 Nov 3;438(7064):57-61. doi: 10.1038/nature04091. Epub 2005 Aug 24. Nature. 2005. PMID: 16121131
-
Emerging non-canonical roles for the Rad51-Rad52 interaction in response to double-strand breaks in yeast.Curr Genet. 2020 Oct;66(5):917-926. doi: 10.1007/s00294-020-01081-z. Epub 2020 May 12. Curr Genet. 2020. PMID: 32399607 Free PMC article. Review.
Cited by
-
Low abundance of telomerase in yeast: implications for telomerase haploinsufficiency.RNA. 2006 Sep;12(9):1721-37. doi: 10.1261/rna.134706. Epub 2006 Aug 7. RNA. 2006. PMID: 16894218 Free PMC article.
-
Defective resection at DNA double-strand breaks leads to de novo telomere formation and enhances gene targeting.PLoS Genet. 2010 May 13;6(5):e1000948. doi: 10.1371/journal.pgen.1000948. PLoS Genet. 2010. PMID: 20485519 Free PMC article.
-
Rapid analysis of Saccharomyces cerevisiae genome rearrangements by multiplex ligation-dependent probe amplification.PLoS Genet. 2012;8(3):e1002539. doi: 10.1371/journal.pgen.1002539. Epub 2012 Mar 1. PLoS Genet. 2012. PMID: 22396658 Free PMC article.
-
The yeast Pif1p DNA helicase preferentially unwinds RNA DNA substrates.Nucleic Acids Res. 2007;35(17):5809-18. doi: 10.1093/nar/gkm613. Epub 2007 Aug 24. Nucleic Acids Res. 2007. PMID: 17720711 Free PMC article.
-
Esc2 promotes telomere stability in response to DNA replication stress.Nucleic Acids Res. 2019 May 21;47(9):4597-4611. doi: 10.1093/nar/gkz158. Nucleic Acids Res. 2019. PMID: 30838410 Free PMC article.
References
-
- Bai Y, Symington LS. A Rad52 homolog is required for RAD51-independent mitotic recombination in Saccharomyces cerevisiae. Genes Dev. 1996;10:2025–2037. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
