

Molecular analysis of the beta-globin gene cluster in the Niokholo Mandenka population reveals a recent origin of the beta(S) Senegal mutation
- PMID: 11741197
- PMCID: PMC384889
- DOI: 10.1086/338304
Molecular analysis of the beta-globin gene cluster in the Niokholo Mandenka population reveals a recent origin of the beta(S) Senegal mutation
Abstract
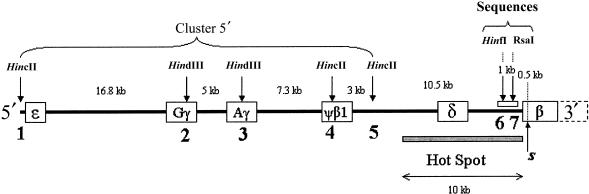
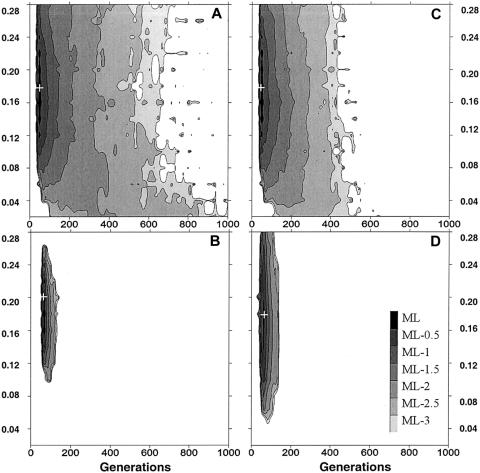
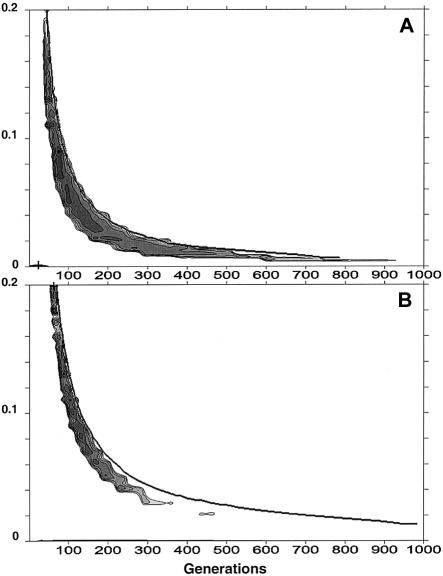
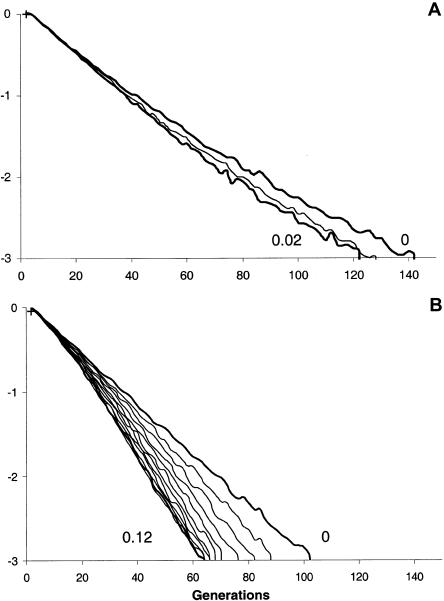
A large and ethnically well-defined Mandenka sample from eastern Senegal was analyzed for the polymorphism of the beta-globin gene cluster on chromosome 11. Five RFLP sites of the 5' region were investigated in 193 individuals revealing the presence of 10 different haplotypes. The frequency of the sickle-cell anemia causing mutation (beta(S)) in the Mandenka estimated from this sample is 11.7%. This mutation was found strictly associated with the single Senegal haplotype. Approximately 600 bp of the upstream region of the beta-globin gene were sequenced for a subset of 94 chromosomes, showing the presence of four transversions, five transitions, and a composite microsatellite polymorphism. The sequence of 22 beta(S) chromosomes was also identical to the previously defined Senegal haplotype, suggesting that this mutation is very recent. Monte Carlo simulations (allowing for a specific balancing selection model, a logistic growth of the population, and variable initial frequencies of the Senegal haplotype) were used to estimate the age of the beta(S) mutation. Resulting maximum-likelihood estimates are 45-70 generations (1,350-2,100 years) for very different demographic scenarios. Smallest confidence intervals (25-690 generations) are obtained under the hypothesis that the Mandenka population is large (N(e) >5,000) and stationary or that it has undergone a rapid demographic expansion to a current size of >5,000 reproducing individuals, which is quite likely in view of the great diversity found on beta(A) chromosomes.
Figures

References
Electronic-Database Information
-
- Bruce Rannala's Research Group, http://www.rannala.org/ (for DMLE and BDMC21)
-
- Statistics, http://www.reading.ac.uk/Statistics/genetics/software.html/ (for hwmetc software)
References
-
- Antonarakis SE, Kazazian HH Jr, Orkin SH (1985) DNA polymorphism and molecular pathology of the human globin gene clusters. Hum Genet 69:1–14 - PubMed
-
- Ayala FJ, Escalante AA, Rich SM (1999) Evolution of Plasmodium and the recent origin of the world populations of Plasmodium falciparum. Parassitologia 41:55–68 - PubMed
-
- Ayres KL, Balding DJ (1998) Measuring departures from Hardy-Weinberg: a Markov chain Monte Carlo method for estimating the inbreeding coefficient. Heredity 80:769–777 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
