

Receptor sensitivity in bacterial chemotaxis
- PMID: 11742065
- PMCID: PMC117525
- DOI: 10.1073/pnas.011589998
Receptor sensitivity in bacterial chemotaxis
Abstract
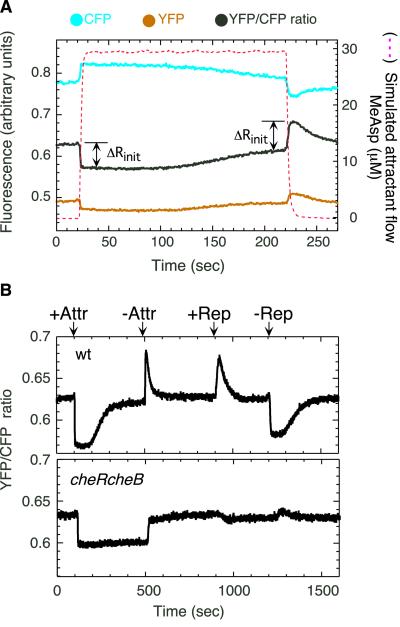
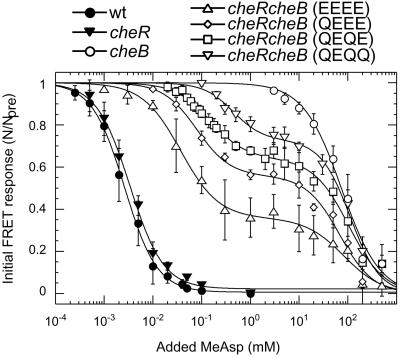
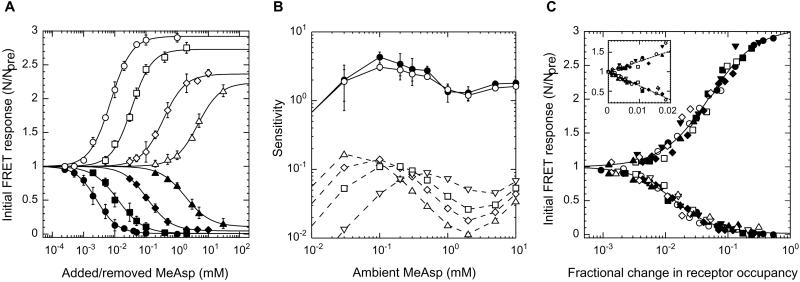
Chemoreceptors in Escherichia coli are coupled to the flagella by a labile phosphorylated intermediate, CheY approximately P. Its activity can be inferred from the rotational bias of flagellar motors, but motor response is stochastic and limited to a narrow physiological range. Here we use fluorescence resonance energy transfer to monitor interactions of CheY approximately P with its phosphatase, CheZ, that reveal changes in the activity of the receptor kinase, CheA, resulting from the addition of attractants or repellents. Analyses of cheR and/or cheB mutants, defective in receptor methylation/demethylation, show that response sensitivity depends on the activity of CheB and the level of receptor modification. In cheRcheB mutants, the concentration of attractant that generates a half-maximal response is equal to the dissociation constant of the receptor. In wild-type cells, it is 35 times smaller. This amplification, together with the ultrasensitivity of the flagellar motor, explains previous observations of high chemotactic gain.
Figures
Comment in
-
Bacterial chemotaxis and the question of gain.Proc Natl Acad Sci U S A. 2002 Jan 8;99(1):7-9. doi: 10.1073/pnas.022641699. Proc Natl Acad Sci U S A. 2002. PMID: 11782543 Free PMC article. No abstract available.
References
-
- Berg H C, Brown D A. Nature (London) 1972;239:500–504. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
