

The gamma -secretase-cleaved C-terminal fragment of amyloid precursor protein mediates signaling to the nucleus
- PMID: 11742091
- PMCID: PMC64969
- DOI: 10.1073/pnas.261463298
The gamma -secretase-cleaved C-terminal fragment of amyloid precursor protein mediates signaling to the nucleus
Abstract
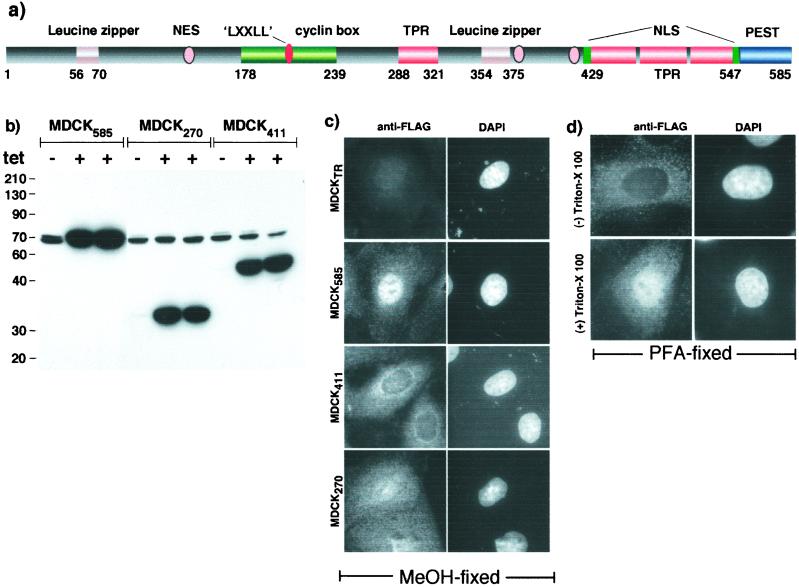
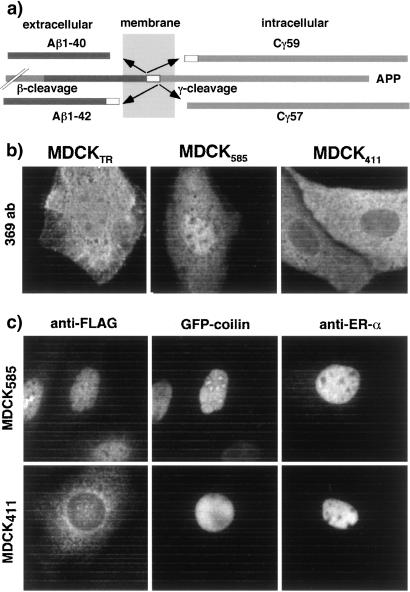
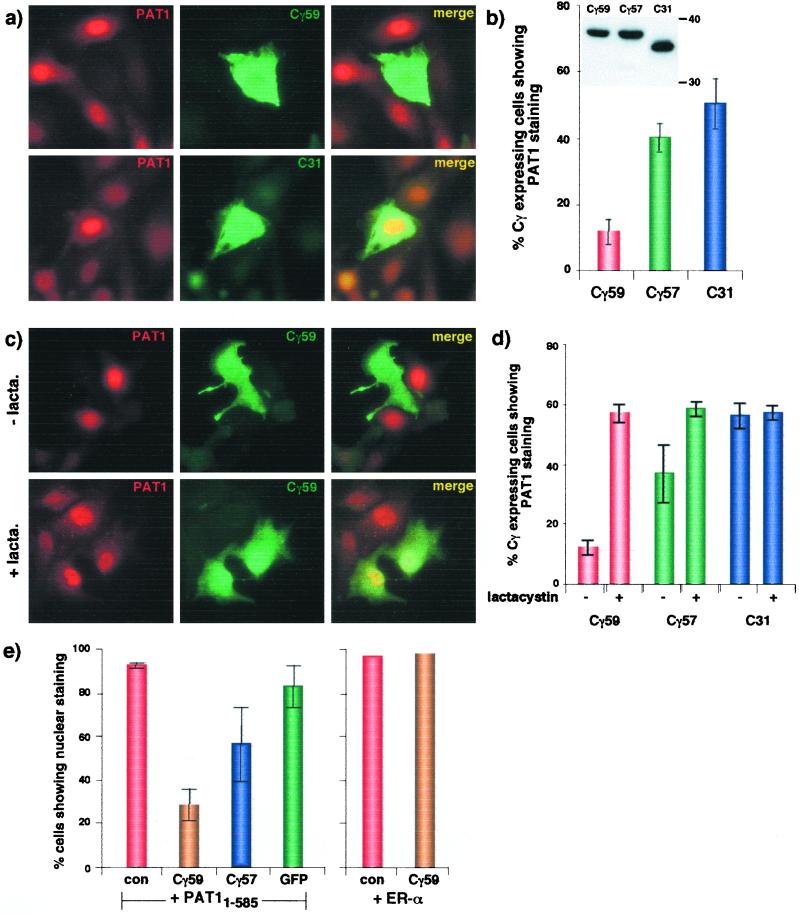
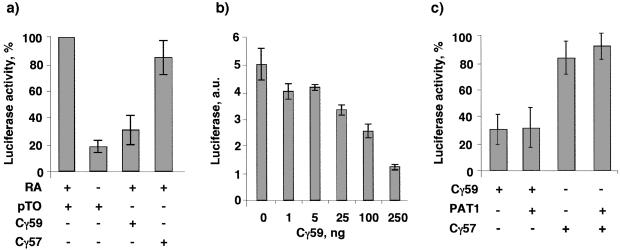
Sequential processing of the amyloid precursor protein (APP) by beta- and gamma-secretases generates the Abeta peptide, a major constituent of the senile plaques observed in Alzheimer's disease. The cleavage by gamma-secretase also results in the cytoplasmic release of a 59- or 57-residue-long C-terminal fragment (Cgamma). This processing resembles regulated intramembrane proteolysis of transmembrane proteins such as Notch, where the released cytoplasmic fragments enter the nucleus and modulate gene expression. Here, we examined whether the analogous Cgamma fragments of APP also exert effects in the nucleus. We find that ectopically expressed Cgamma is present both in the cytoplasm and in the nucleus. Interestingly, expression of Cgamma59 causes disappearance of PAT1, a protein that interacts with the APP cytoplasmic domain, from the nucleus and induces its proteosomal degradation. Treatment of cells with lactacystin prevents PAT1 degradation and retains its nuclear localization. By contrast, Cgamma57, a minor product of gamma-cleavage, is only marginally effective in PAT1 degradation. Furthermore, Cgamma59 but not Cgamma57 potently represses retinoic acid-responsive gene expression. Thus, our studies provide the evidence that, as predicted by the regulated intramembrane proteolysis mechanism, Cgamma seems to function in the nucleus.
Figures
References
-
- Price D L, Sisodia S S. Annu Rev Neurosci. 1998;21:479–505. - PubMed
-
- Selkoe D J. Trends Cell Biol. 1998;8:447–453. - PubMed
-
- De Strooper B, Annaert W. J Cell Sci. 2000;113:1857–1870. - PubMed
-
- Vassar R, Citron M. Neuron. 2000;27:419–422. - PubMed
-
- Haass C, De Strooper B. Science. 1999;286:916–919. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
