

Cell-to-cell movement and assembly of a plant closterovirus: roles for the capsid proteins and Hsp70 homolog
- PMID: 11742977
- PMCID: PMC125784
- DOI: 10.1093/emboj/20.24.6997
Cell-to-cell movement and assembly of a plant closterovirus: roles for the capsid proteins and Hsp70 homolog
Abstract
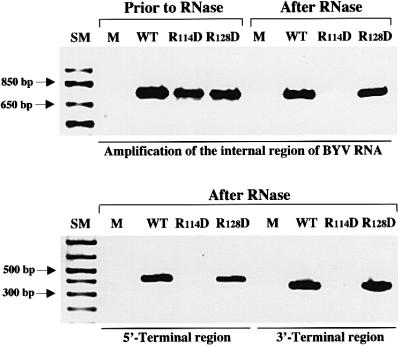
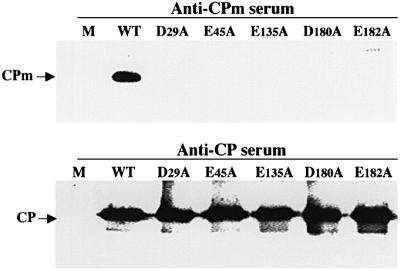
Diverse animal and plant viruses are able to translocate their virions between neighboring cells via intercellular connections. In this work, we analyze the virion assembly and cell-to-cell movement of a plant closterovirus and reveal a strong correlation between these two processes. The filamentous virions of a closterovirus possess a long body formed by the major capsid protein (CP) and a short tail formed by the minor capsid protein (CPm). Genetic and biochemical analyses show that the functions of these virion components are distinct. A virion body is required primarily for genome protection, whereas a tail represents a specialized device for cell-to-cell movement. Furthermore, tail assembly is mediated by the viral Hsp70 homolog (Hsp70h) that becomes an integral part of the virion. Inactivation of the ATPase domain of Hsp70h results in assembly of tailless virions that are incapable of translocation. A dual role for the viral molecular chaperone Hsp70h in virion assembly and transport, combined with the previous finding of this protein in intercellular channels, allowed us to propose a model of closteroviral movement from cell to cell.
Figures








References
-
- Agranovsky A.A., Boyko,V.P., Karasev,A.V., Koonin,E.V. and Dolja,V.V. (1991) Putative 65 kDa protein of beet yellows closterovirus is a homologue of HSP70 heat shock proteins. J. Mol. Biol., 217, 603–610. - PubMed
-
- Alzhanova D.V., Hagiwara,Y., Peremyslov,V.V. and Dolja,V.V. (2000) Genetic analysis of the cell-to-cell movement of beet yellows closterovirus. Virology, 267, 192–200. - PubMed
-
- Boorstein W.R., Ziegelhoffer,T. and Craig,E.A. (1994) Molecular evolution of the HSP70 multigene family. J. Mol. Evol., 38, 1–17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
