

Phosphorylation of Nedd4-2 by Sgk1 regulates epithelial Na(+) channel cell surface expression
- PMID: 11742982
- PMCID: PMC125341
- DOI: 10.1093/emboj/20.24.7052
Phosphorylation of Nedd4-2 by Sgk1 regulates epithelial Na(+) channel cell surface expression
Abstract
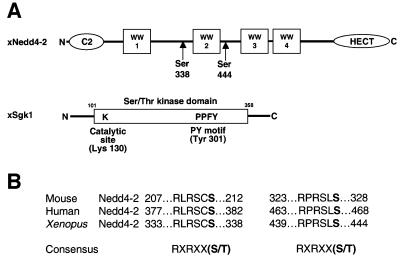
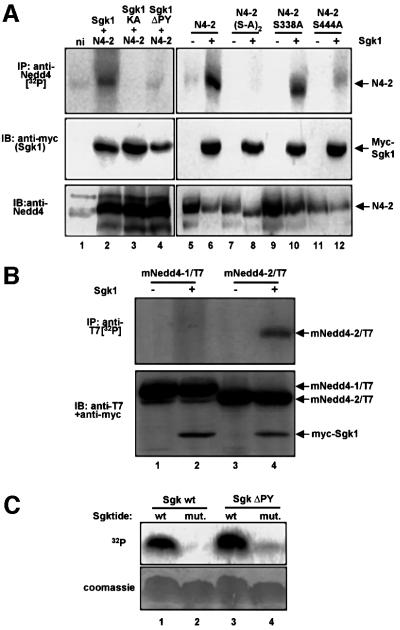
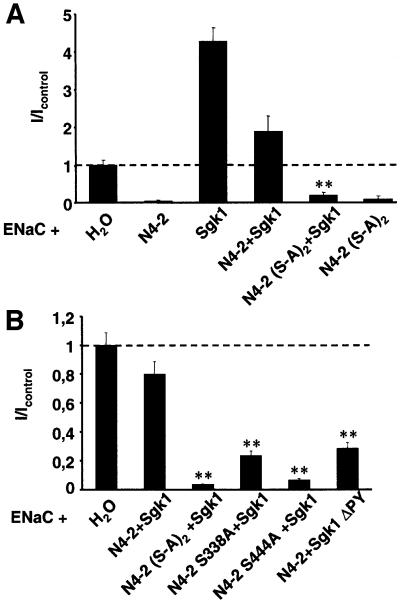
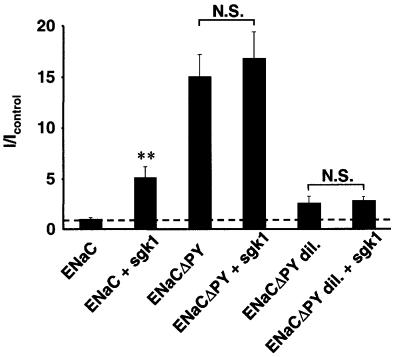
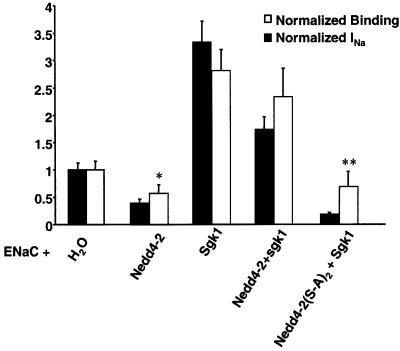
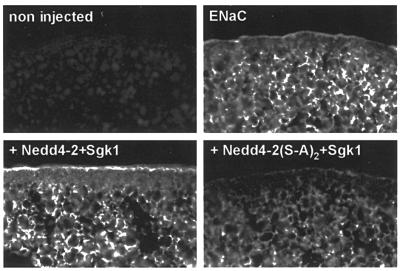
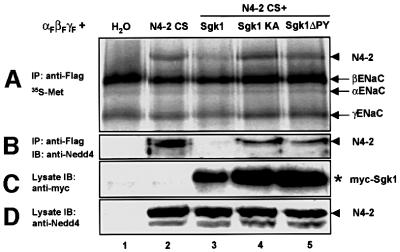
The epithelial Na(+) channel (ENaC) plays an essential role in the regulation of whole body Na(+) balance and blood pressure. The cell surface expression of this channel, a complex of three subunits (alpha, beta and gamma ENaC), has been shown to be regulated by hormones such as aldosterone and vasopressin and by intracellular signaling, including ubiquitylation and/or phosphorylation. However, the molecular mechanisms involving phosphorylation in the regulation of ENaC are unclear. Here we show by expression studies in Xenopus laevis oocytes that the aldosterone-induced Sgk1 kinase interacts with the ubiquitin protein ligase Nedd4-2 in a PY motif-dependent manner and phosphorylates Nedd4-2 on Ser444 and, to a lesser extent, Ser338. Such phosphorylation reduces the interaction between Nedd4-2 and ENaC, leading to elevated ENaC cell surface expression. These data show that phosphorylation of an enzyme involved in the ubiquitylation cascade (Nedd4-2) controls cell surface density of ENaC and propose a paradigm for the control of ion channels. Moreover, they suggest a novel and complete signaling cascade for aldosterone-dependent regulation of ENaC.
Figures
References
-
- Abriel H., Kamynina,E., Horisberger,J.-D. and Staub,O. (2000) Regulation of the cardiac voltage-gated Na+ channel (rH1) by the ubiquitin-protein ligase Nedd4. FEBS Lett., 466, 377–380. - PubMed
-
- Alvarez de la Rosa D., Zhang,P., Naray-Fejes-Toth,A., Fejes-Toth,G. and Canessa,C.M. (1999) The serum and glucocorticoid kinase sgk increases the abundance of epithelial sodium channels in the plasma membrane of Xenopus oocytes. J. Biol. Chem., 274, 37834–37839. - PubMed
-
- Blazer-Yost B.L., Paunescu,T.G., Helman,S.I., Lee,K.D. and Vlahos,C.J. (1999) Phosphoinositide 3-kinase is required for aldosterone-regulated sodium reabsorption. Am. J. Physiol., 277, C531–C536. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
