

Flipping a genetic switch by subunit exchange
- PMID: 11742991
- PMCID: PMC125793
- DOI: 10.1093/emboj/20.24.7149
Flipping a genetic switch by subunit exchange
Erratum in
- EMBO J. 2004 Aug 4;23(15):3186
Abstract
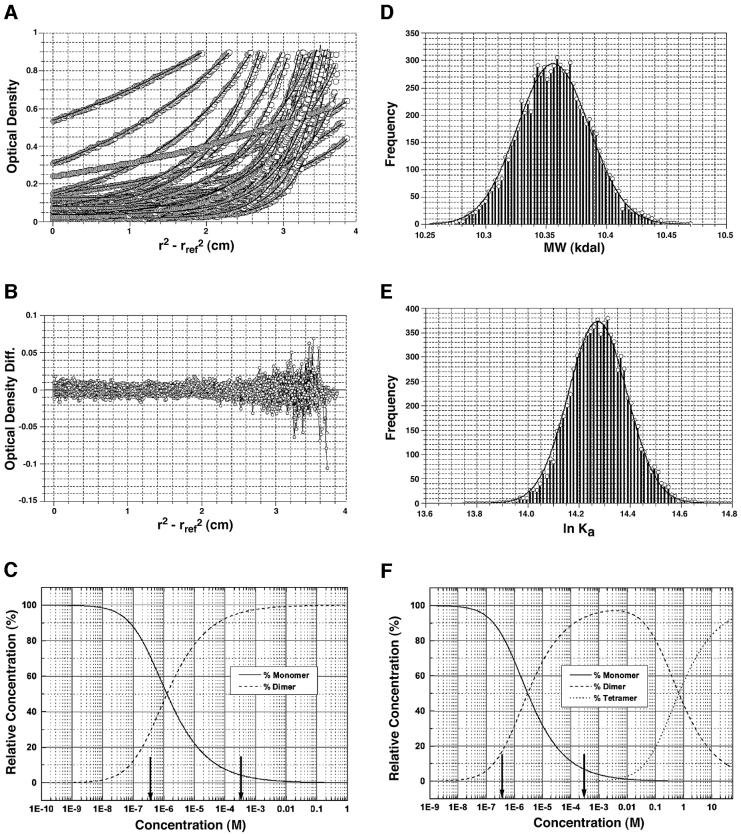
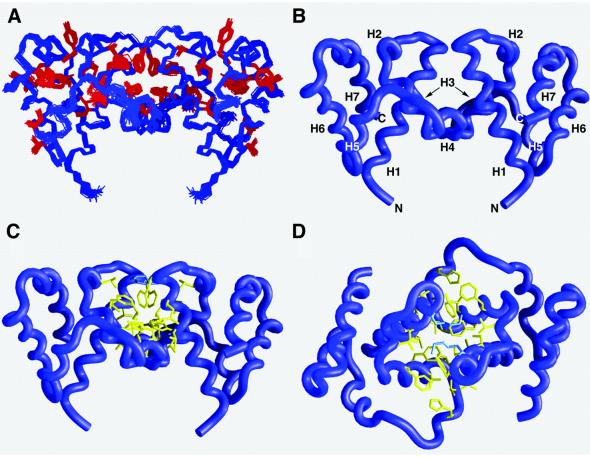
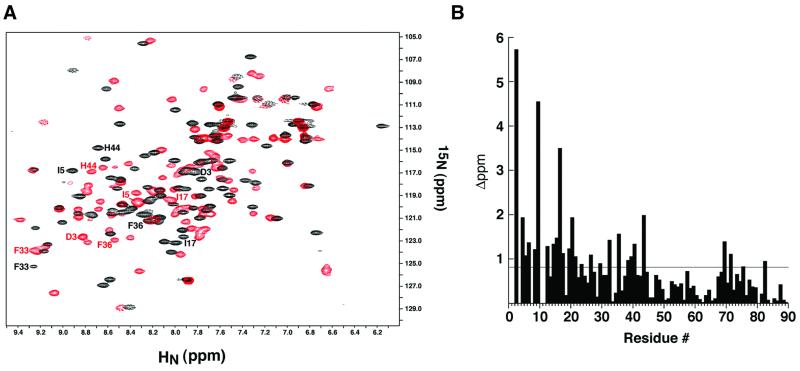
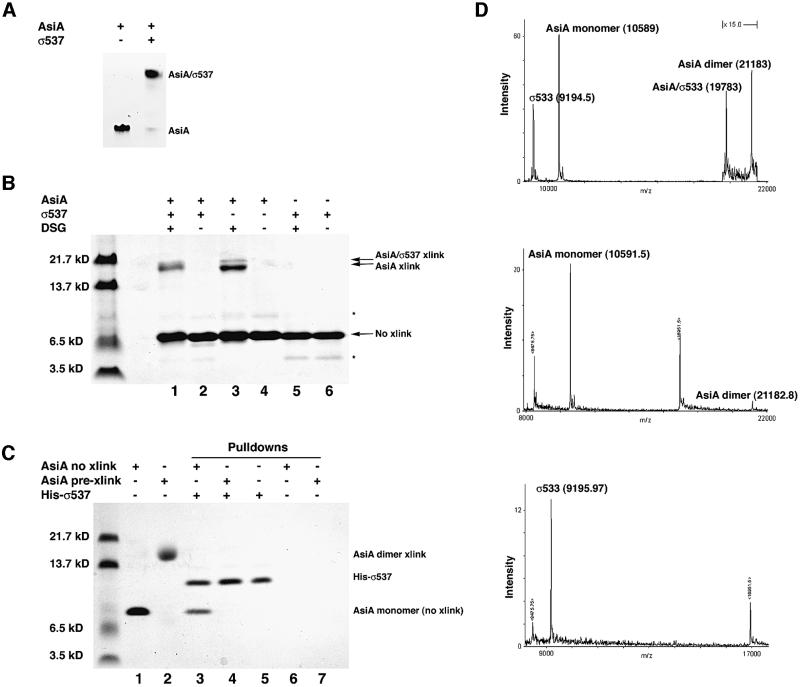
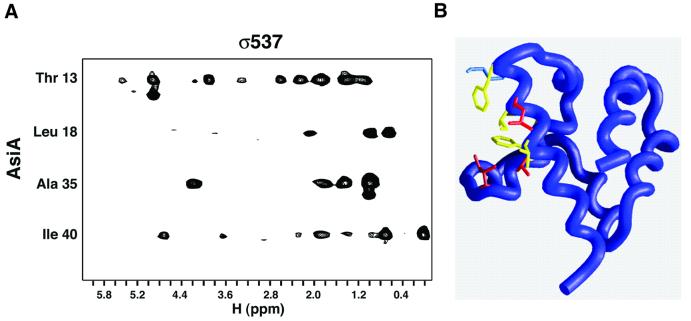
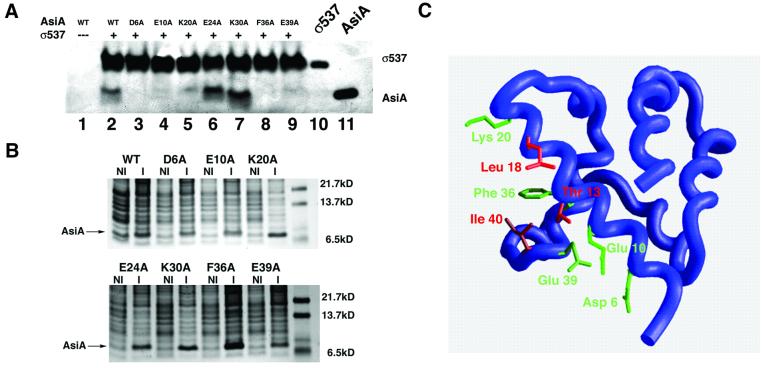
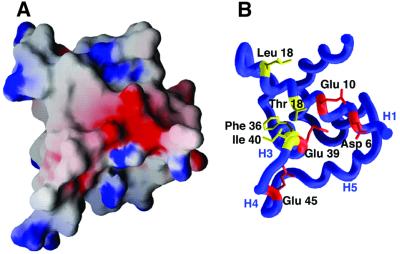
The bacteriophage T4 AsiA protein is a multifunctional protein that simultaneously acts as both a repressor and activator of gene expression during the phage life cycle. These dual roles with opposing transcriptional consequences are achieved by modification of the host RNA polymerase in which AsiA binds to conserved region 4 (SR4) of sigma(70), altering the pathway of promoter selection by the holoenzyme. The mechanism by which AsiA flips this genetic switch has now been revealed, in part, from the three-dimensional structure of AsiA and the elucidation of its interaction with SR4. The structure of AsiA is that of a novel homodimer in which each monomer is constructed as a seven-helix bundle arranged in four overlapping helix-loop-helix elements. Identification of the protein interfaces for both the AsiA homodimer and the AsiA-sigma(70) complex reveals that these interfaces are coincident. Thus, the AsiA interaction with sigma(70) necessitates that the AsiA homodimer dissociate to form an AsiA-SR4 heterodimer, exchanging one protein subunit for another to alter promoter choice by RNA polymerase.
Figures
References
-
- Adelman K., Orsini,G., Kolb,A., Graziani,L. and Brody,E.N. (1997) The interaction between the AsiA protein of bacteriophage T4 and the σ70 subunit of Escherichia coli RNA polymerase. J. Biol. Chem., 272, 27435–27443. - PubMed
-
- Brünger A.T. (1992) X-PLOR Manual, Version 3.1. Yale University Press, New Haven, CT.
-
- Bushman F.D., Shang,C. and Ptashne,M. (1989) A single glutamic acid residue plays a key role in the transcriptional activation function of λ repressor. Cell, 58, 1163–1171. - PubMed
-
- Cadene M. and Chait,B.T. (2000) A robust, detergent-friendly method for mass spectrometric analysis of integral membrane proteins. Anal. Chem., 72, 5655–5658. - PubMed
-
- Campbell E.A. and Darst,S.A. (2000) The anti-σ factor SpoIIAB forms a 2:1 complex with σF, contacting multiple conserved regions of the σ factor. J. Mol. Biol., 300, 17–28. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
