

Repression of inflammatory responses in the absence of DNA binding by the glucocorticoid receptor
- PMID: 11742993
- PMCID: PMC125338
- DOI: 10.1093/emboj/20.24.7168
Repression of inflammatory responses in the absence of DNA binding by the glucocorticoid receptor
Abstract
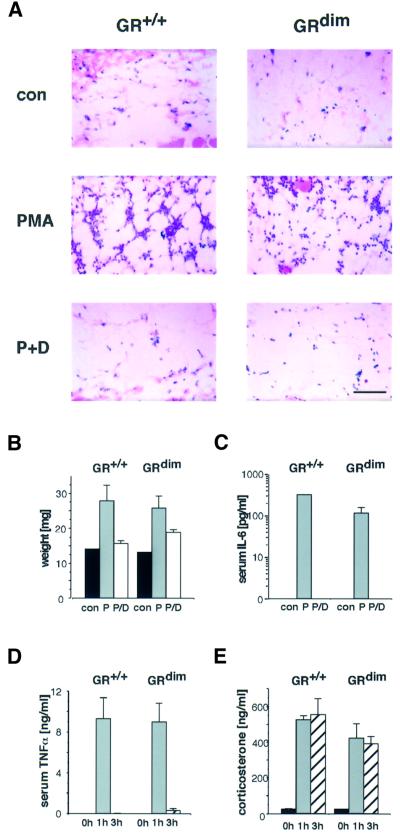
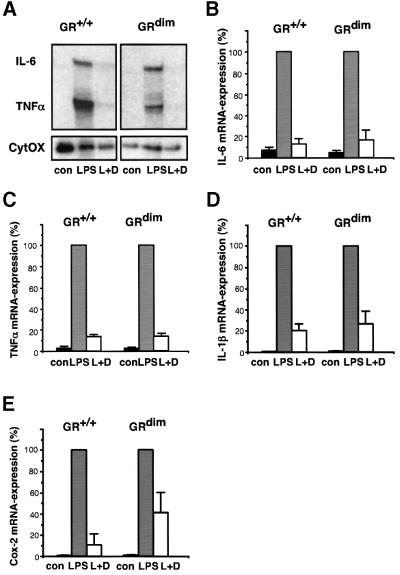
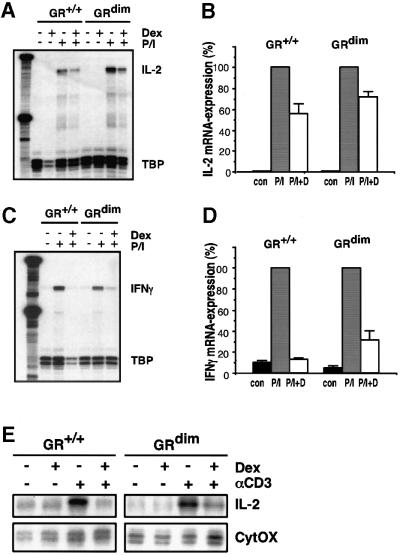
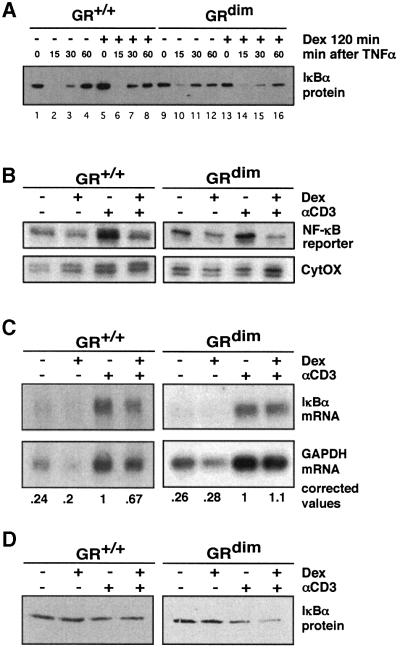
The glucocorticoid receptor (GR) acts both as a transcription factor itself on genes carrying GR response elements (GREs) and as a modulator of other transcription factors. Using mice with a mutation in the GR, which cannot activate GRE promoters, we examine whether the important anti-inflammatory and immune suppressive functions of glucocorticoids (GCs) can be established in this in vivo animal model. We find that most actions are indeed exerted in the absence of the DNA-binding ability of the GR: inhibition of the inflammatory response of locally irritated skin and of the systemic response to lipopolysaccharides. GCs repress the expression and release of numerous cytokines both in vivo and in isolated primary macrophages, thymocytes and CD4(+) splenocytes. A transgenic reporter gene controlled by NF-kappa B exclusively is also repressed, suggesting that protein- protein interaction with other transcription factors such as NF-kappa B forms the basis of the anti-inflammatory activity of GR. The only defect of immune suppression detected so far concerns the induced apoptosis of thymocytes and T lymphocytes.
Figures
References
-
- Auphan N., DiDonato,J.A., Rosette,C., Helmberg,A. and Karin,M. (1995) Immunosuppression by glucocorticoids: inhibition of NF-κB activity through induction of IκB synthesis. Science, 270, 286–290. - PubMed
-
- Baeuerle P.A. and Baltimore,D. (1996) NF-κB: ten years after. Cell, 87, 13–20. - PubMed
-
- Barnes P.J. (1998) Anti-inflammatory actions of glucocorticoids: molecular mechanisms. Clin. Sci., 94, 557–572. - PubMed
-
- Beato M., Herrlich,P. and Schütz,G. (1995) Steroid hormone receptors: many actors in search of a plot. Cell, 83, 851–857. - PubMed
-
- Brostjan C., Anrather,J., Csizmadia,V., Natarajan,G. and Winkler,H. (1997) Glucocorticoids inhibit E-selectin expression by targeting NF-κB and not ATF/c-Jun. J. Immunol., 158, 3836–3844. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
