

The RecOR proteins modulate RecA protein function at 5' ends of single-stranded DNA
- PMID: 11743007
- PMCID: PMC125792
- DOI: 10.1093/emboj/20.24.7313
The RecOR proteins modulate RecA protein function at 5' ends of single-stranded DNA
Abstract
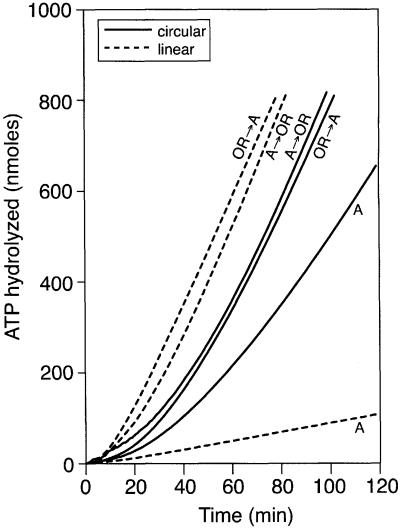
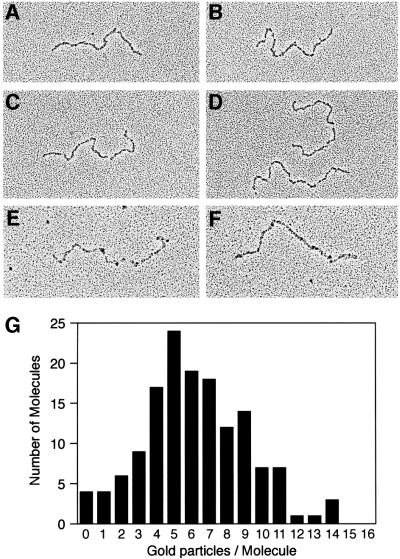
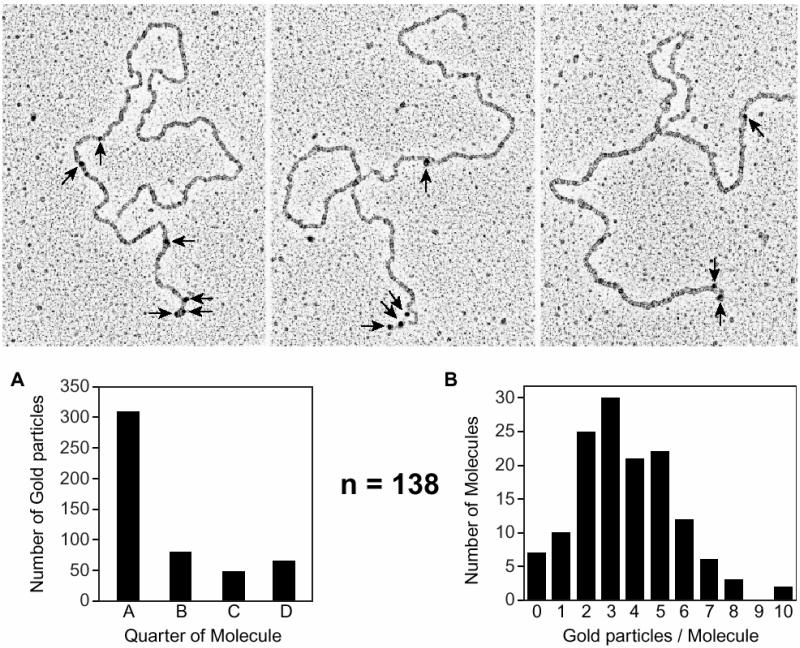
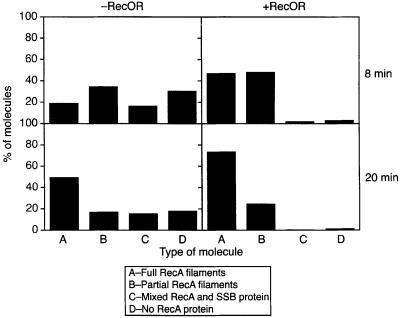
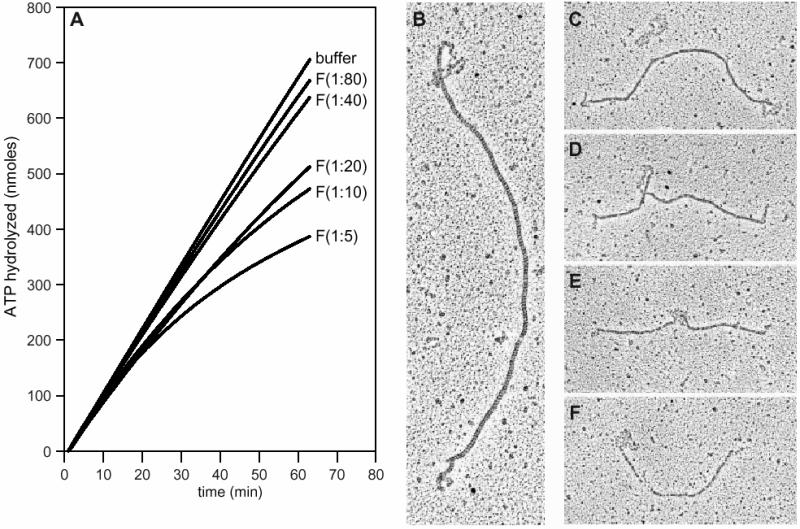
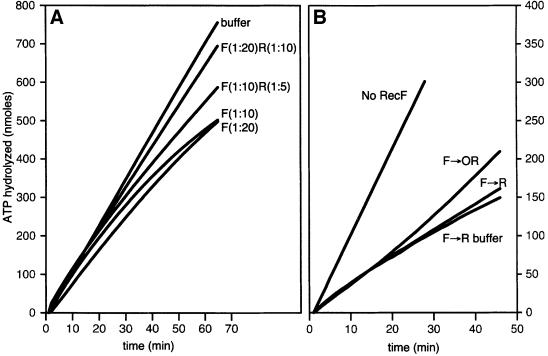
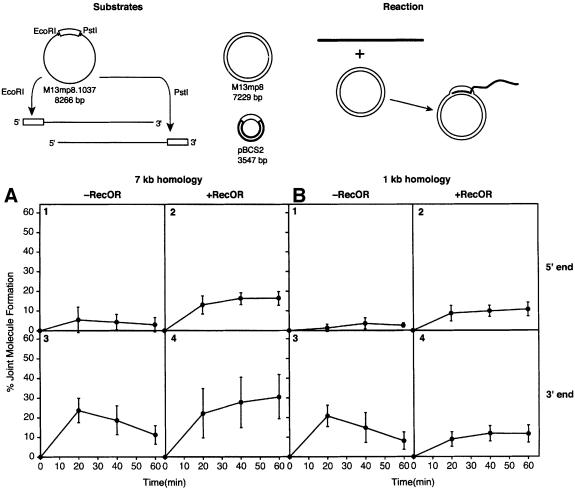
The Escherichia coli RecF, RecO and RecR pro teins have previously been implicated in bacterial recombinational DNA repair at DNA gaps. The RecOR-facilitated binding of RecA protein to single-stranded DNA (ssDNA) that is bound by single-stranded DNA-binding protein (SSB) is much faster if the ssDNA is linear, suggesting that a DNA end (rather than a gap) facilitates binding. In addition, the RecOR complex facilitates RecA protein-mediated D-loop formation at the 5' ends of linear ssDNAs. RecR protein remains associated with the RecA filament and its continued presence is required to prevent filament disassembly. RecF protein competes with RecO protein for RecR protein association and its addition destabilizes RecAOR filaments. An enhanced function of the RecO and RecR proteins can thus be seen in vitro at the 5' ends of linear ssDNA that is not as evident in DNA gaps. This function is countered by the RecF/RecO competition for association with the RecR protein.
Figures
References
-
- Beernink H.T.H. and Morrical,S.W. (1999) RMPs: recombination/replication mediator proteins. Trends Biochem. Sci., 24, 385–389. - PubMed
-
- Bork J.M., Cox,M.M. and Inman,R.B. (2001) RecA protein filaments disassemble in the 5′ to 3′ direction on single-stranded DNA. J. Biol. Chem., 276, in press. - PubMed
-
- Bradford M.M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem., 72, 248–254. - PubMed
-
- Clark A.J. and Sandler,S.J. (1994) Homologous genetic recombination: the pieces begin to fall into place. Crit. Rev. Microbiol., 20, 125–142. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
