

Ionic strength- and temperature-induced K(Ca) shifts in the uncoating reaction of rotavirus strains RF and SA11: correlation with membrane permeabilization
- PMID: 11752146
- PMCID: PMC136821
- DOI: 10.1128/jvi.76.2.552-559.2002
Ionic strength- and temperature-induced K(Ca) shifts in the uncoating reaction of rotavirus strains RF and SA11: correlation with membrane permeabilization
Abstract
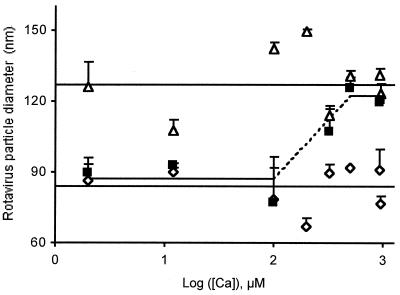
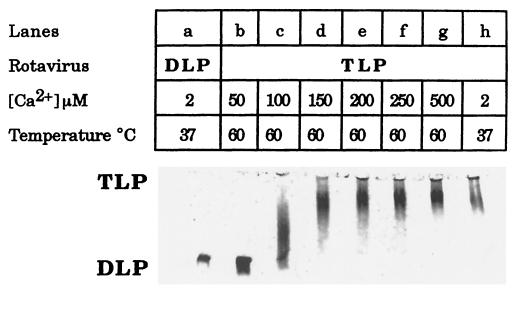
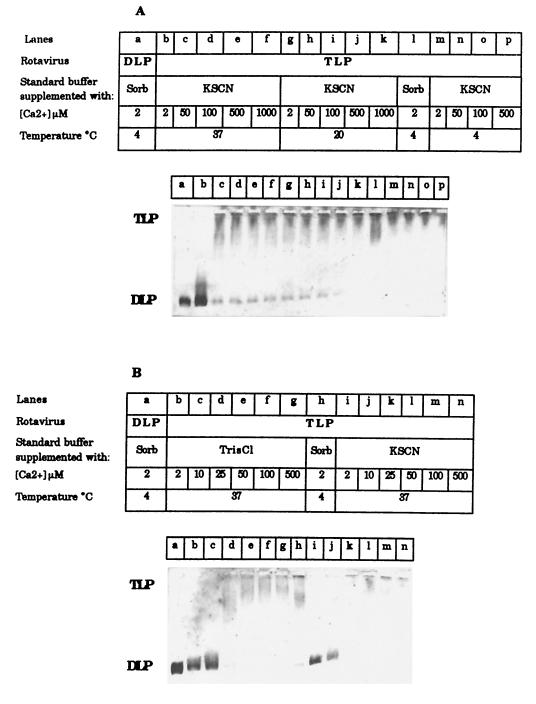
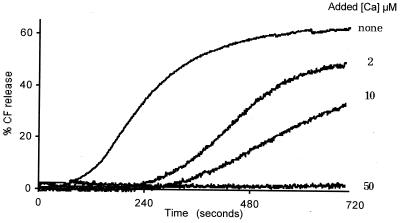
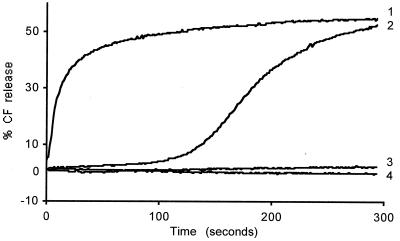
The hydrodynamic diameters of native rotavirus particles, bovine RF and simian SA11 strains, were determined by quasielastic light scattering. By using this method and agarose gel electrophoresis, the Ca(2+) dissociation constant, K(Ca), governing the transition from triple-layer particles (TLPs) to double-layer particles (DLPs), was shown to increase, at constant pH, as the temperature and/or the ionic strength of the incubation medium increased. We report the novel observation that, under physiological conditions, K(Ca) values for both RF and SA11 rotaviruses were well above the intracytoplasmic Ca(2+) concentrations of various cells, which may explain why TLP uncoating takes place within vesicles (possibly endosomes) during the entry process. A correlation between TLP uncoating and cell membrane permeabilization was found, as shown by the release of carboxyfluorescein (CF) from CF-loaded intestinal brush-border membrane vesicles. Conditions stabilizing the virion in the TLP form inhibited CF release, whereas conditions favoring the TLP-to-DLP transformation activated this process. We conclude that membrane permeabilization must be preceded by the loss of the outer-capsid proteins from trypsinized TLP and that physiological ionic strength is required for permeabilization to take place. Finally, the paper develops an alternative explanation for the mechanism of rotavirus entry, compatible with the Ca(2+)-dependent endocytic pathway. We propose that there must be an iterative process involving tight coupling in time between the lowering of endosomal Ca(2+) concentration, virion decapsidation, and membrane permeabilization, which would cause the transcriptionally active DLPs to enter the cytoplasm of cells.
Figures

Similar articles
-
Visualization of Calcium Ion Loss from Rotavirus during Cell Entry.J Virol. 2018 Nov 27;92(24):e01327-18. doi: 10.1128/JVI.01327-18. Print 2018 Dec 15. J Virol. 2018. PMID: 30258012 Free PMC article.
-
Rotavirus interaction with isolated membrane vesicles.J Virol. 1994 Jun;68(6):4009-16. doi: 10.1128/JVI.68.6.4009-4016.1994. J Virol. 1994. PMID: 8189534 Free PMC article.
-
Cell lines susceptible to infection are permeabilized by cleaved and solubilized outer layer proteins of rotavirus.J Gen Virol. 1997 Nov;78 ( Pt 11):2883-93. doi: 10.1099/0022-1317-78-11-2883. J Gen Virol. 1997. PMID: 9367375
-
Signal transduction and ion channels in guard cells.Philos Trans R Soc Lond B Biol Sci. 1998 Sep 29;353(1374):1475-88. doi: 10.1098/rstb.1998.0303. Philos Trans R Soc Lond B Biol Sci. 1998. PMID: 9800209 Free PMC article. Review.
-
Cytoplasmic zoning by protein phase transition after membrane permeabilization.J Biochem. 2024 Feb 25;175(2):147-153. doi: 10.1093/jb/mvad094. J Biochem. 2024. PMID: 37972304 Free PMC article. Review.
Cited by
-
Cross-linking of rotavirus outer capsid protein VP7 by antibodies or disulfides inhibits viral entry.J Virol. 2011 Oct;85(20):10509-17. doi: 10.1128/JVI.00234-11. Epub 2011 Aug 17. J Virol. 2011. PMID: 21849465 Free PMC article.
-
Molecular basis for the different PCV2 susceptibility of T-lymphoblasts in Landrace and Piétrain pigs.Vet Res. 2024 Feb 19;55(1):22. doi: 10.1186/s13567-024-01275-0. Vet Res. 2024. PMID: 38374131 Free PMC article.
-
Solar and temperature treatments affect the ability of human rotavirus wa to bind to host cells and synthesize viral RNA.Appl Environ Microbiol. 2015 Jun 15;81(12):4090-7. doi: 10.1128/AEM.00027-15. Epub 2015 Apr 10. Appl Environ Microbiol. 2015. PMID: 25862222 Free PMC article.
-
VP5* rearranges when rotavirus uncoats.J Virol. 2009 Nov;83(21):11372-7. doi: 10.1128/JVI.01228-09. Epub 2009 Aug 19. J Virol. 2009. PMID: 19692464 Free PMC article.
-
Adenovirus protein VI mediates membrane disruption following capsid disassembly.J Virol. 2005 Feb;79(4):1992-2000. doi: 10.1128/JVI.79.4.1992-2000.2005. J Virol. 2005. PMID: 15681401 Free PMC article.
References
-
- Charpilienne, A., M. J. Abad, F. Michelangeli, F. Alvarado, M. Vasseur, J. Cohen, and M. C. Ruiz. 1997. Solubilized and cleaved VP7, the outer glycoprotein of rotavirus, induces permeabilization of cell membrane vesicles. J. Gen. Virol. 78:1367–1371. - PubMed
-
- Cohen, J., J. Laporte, A. Charpilienne, and R. Scherrer. 1979. Activation of rotavirus RNA polymerase by calcium chelation. Arch. Virol. 60:177–186. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
