

The pseudorabies virus VP22 homologue (UL49) is dispensable for virus growth in vitro and has no effect on virulence and neuronal spread in rodents
- PMID: 11752167
- PMCID: PMC136827
- DOI: 10.1128/jvi.76.2.774-782.2002
The pseudorabies virus VP22 homologue (UL49) is dispensable for virus growth in vitro and has no effect on virulence and neuronal spread in rodents
Abstract
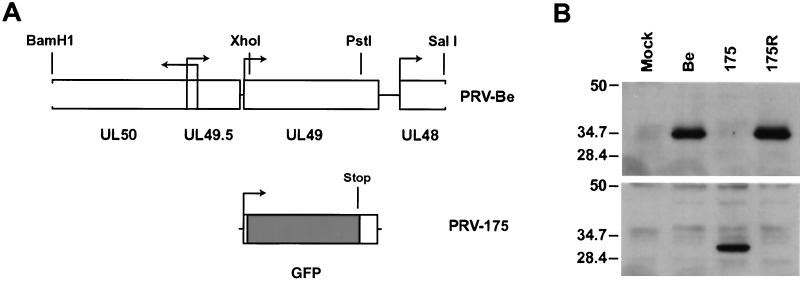
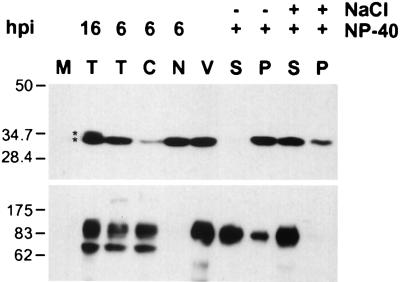
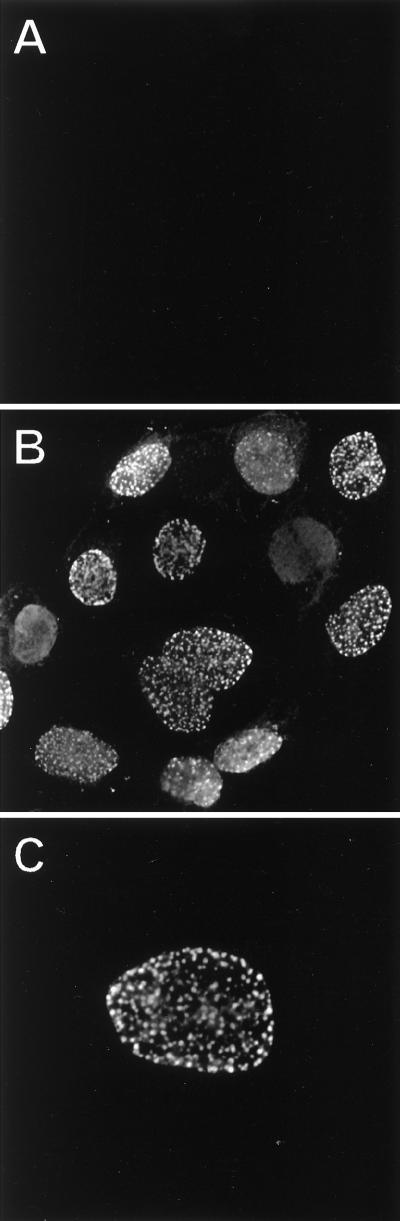
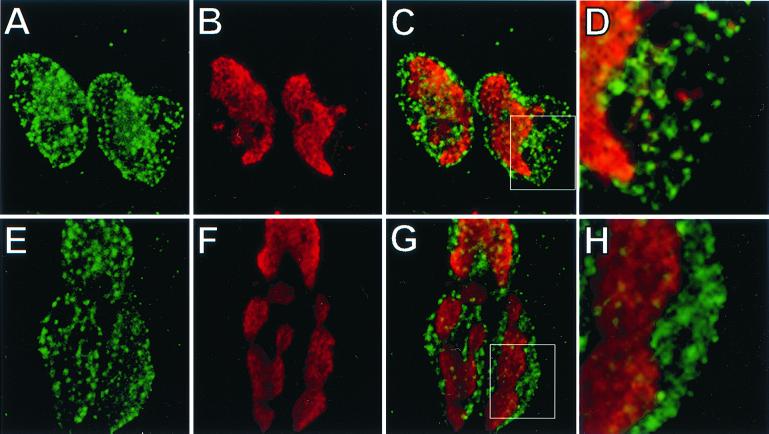
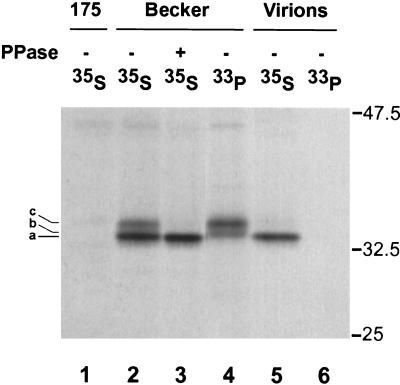
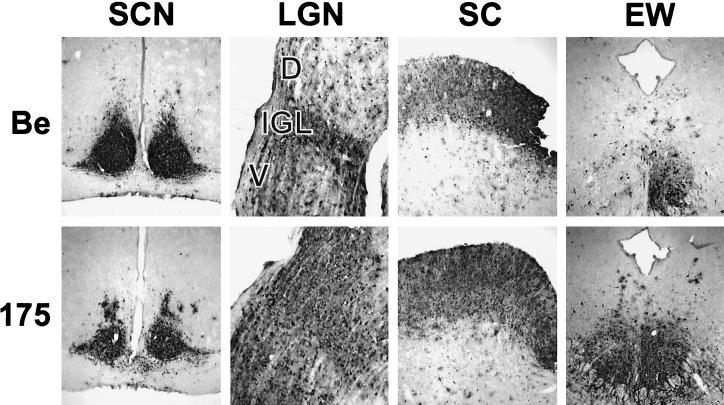
The tegument of herpesvirus virions is a distinctive structure whose assembly and function are not well understood. The herpes simplex virus type 1 VP22 tegument protein encoded by the UL49 gene is conserved among the alphaherpesviruses. Using cell biology and viral genetics, we provide an initial characterization of the pseudorabies virus (PRV) VP22 homologue. We identified three isoforms of VP22 present in PRV-infected cells that can be resolved by polyacrylamide gel electrophoresis. The predominant form is not phosphorylated and is present in virions, while the other two species are phosphorylated and excluded from virions. VP22 localized to the nucleus by 6 h postinfection, as determined by immunofluorescence and cell fractionation. VP22 immunofluorescence in the nucleus was both diffuse and in punctate structures. The punctate nuclear localization was the most pronounced form of staining and did not localize exclusively to sites of viral DNA replication. Unexpectedly, a VP22 null mutant had no obvious phenotypes during tissue culture infections and was similar to the wild type in all respects. Moreover, the VP22 null mutant was as virulent and neuroinvasive as the wild-type virus after infection of the rodent eye and spread to the brain using both anterograde and retrograde neuronal circuits.
Figures
Similar articles
-
[Identification of nuclear localization signals of pseudorabies virus gene UL49].Bing Du Xue Bao. 2014 Jul;30(4):436-40. Bing Du Xue Bao. 2014. PMID: 25272600 Chinese.
-
Actin is a component of the compensation mechanism in pseudorabies virus virions lacking the major tegument protein VP22.J Virol. 2005 Jul;79(13):8614-9. doi: 10.1128/JVI.79.13.8614-8619.2005. J Virol. 2005. PMID: 15956602 Free PMC article.
-
The UL48 tegument protein of pseudorabies virus is critical for intracytoplasmic assembly of infectious virions.J Virol. 2002 Jul;76(13):6729-42. doi: 10.1128/jvi.76.13.6729-6742.2002. J Virol. 2002. PMID: 12050386 Free PMC article.
-
Molecular biology of pseudorabies virus: impact on neurovirology and veterinary medicine.Microbiol Mol Biol Rev. 2005 Sep;69(3):462-500. doi: 10.1128/MMBR.69.3.462-500.2005. Microbiol Mol Biol Rev. 2005. PMID: 16148307 Free PMC article. Review.
-
Pseudorabies virus neuroinvasiveness: a window into the functional organization of the brain.Adv Virus Res. 2001;56:39-71. doi: 10.1016/s0065-3527(01)56004-2. Adv Virus Res. 2001. PMID: 11450308 Review. No abstract available.
Cited by
-
Characterization of VP22 in herpes simplex virus-infected cells.J Virol. 2005 Oct;79(19):12185-98. doi: 10.1128/JVI.79.19.12185-12198.2005. J Virol. 2005. PMID: 16160145 Free PMC article.
-
The capsid and tegument of the alphaherpesviruses are linked by an interaction between the UL25 and VP1/2 proteins.J Virol. 2007 Nov;81(21):11790-7. doi: 10.1128/JVI.01113-07. Epub 2007 Aug 22. J Virol. 2007. PMID: 17715218 Free PMC article.
-
Nuclear sequestration of cellular chaperone and proteasomal machinery during herpes simplex virus type 1 infection.J Virol. 2004 Jul;78(13):7175-85. doi: 10.1128/JVI.78.13.7175-7185.2004. J Virol. 2004. PMID: 15194794 Free PMC article.
-
Functions of the ORF9-to-ORF12 gene cluster in varicella-zoster virus replication and in the pathogenesis of skin infection.J Virol. 2008 Jun;82(12):5825-34. doi: 10.1128/JVI.00303-08. Epub 2008 Apr 9. J Virol. 2008. PMID: 18400847 Free PMC article.
-
Herpes simplex virus glycoproteins gD and gE/gI serve essential but redundant functions during acquisition of the virion envelope in the cytoplasm.J Virol. 2003 Aug;77(15):8481-94. doi: 10.1128/jvi.77.15.8481-8494.2003. J Virol. 2003. PMID: 12857917 Free PMC article.
References
-
- Ben-Porat, T., and A. S. Kaplan. 1985. Molecular biology of pseudorabies virus, p.105–173. In B. Roizman (ed.), The herpesviruses. Plenum Publishing, New York, N.Y.
-
- Bresnahan, W. A., and T. Shenk. 2000. A subset of viral transcripts packaged within human cytomegalovirus particles. Science 288:2373–2376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
