

The double par locus of virulence factor pB171: DNA segregation is correlated with oscillation of ParA
- PMID: 11752455
- PMCID: PMC64986
- DOI: 10.1073/pnas.261569598
The double par locus of virulence factor pB171: DNA segregation is correlated with oscillation of ParA
Abstract
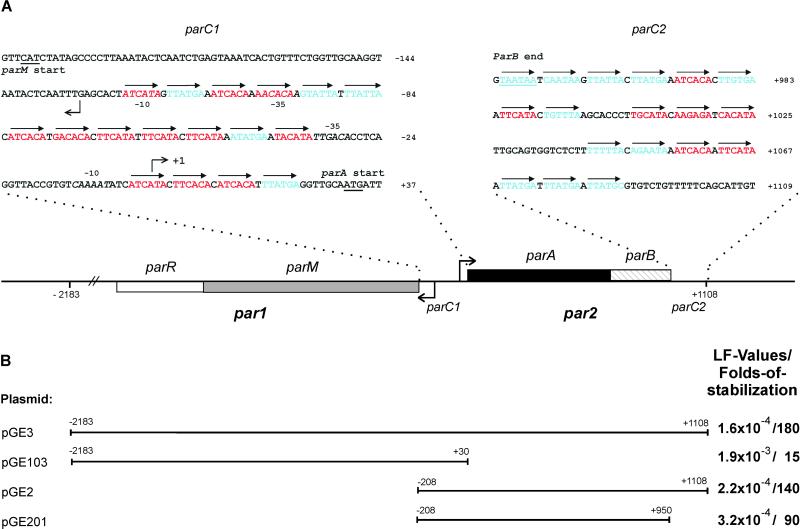
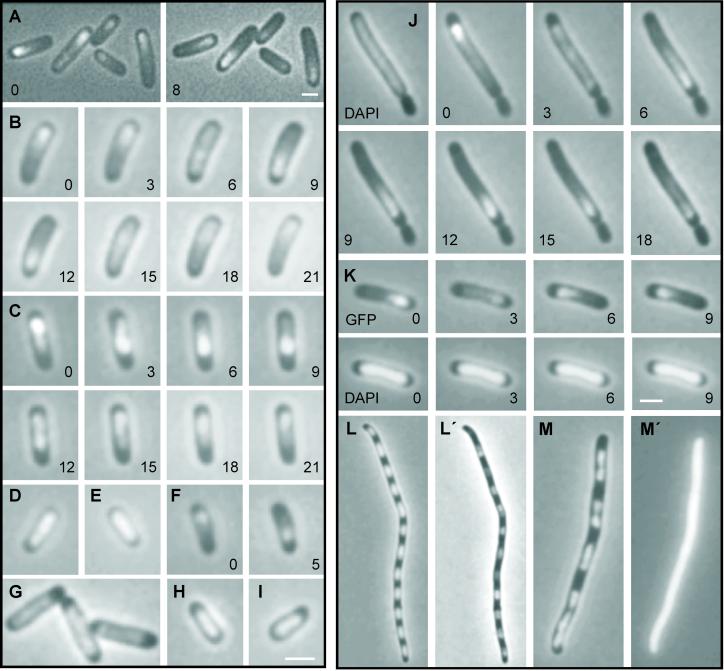
Prokaryotic plasmids and chromosomes encode partitioning (par) loci that segregate DNA to daughter cells before cell division. Recent database analyses showed that almost all known par loci encode an ATPase and a DNA-binding protein, and one or more cis-acting regions where the proteins act. All par-encoded ATPases belong to one of two protein superfamilies, Walker-type and actin-like ATPases. This property was recently used to divide par loci into Types I and II loci. We show here that the Escherichia coli virulence factor pB171 encodes a double par locus that consists of one Type I and one Type II locus. Separately, each locus stabilized a test-plasmid efficiently. Together, the two loci mediated even more efficient plasmid stabilization. The par loci have a unique genetic organization in that they share a common central region at which the two different DNA-binding proteins probably act. Interestingly, a fusion protein consisting of the Walker-type ParA ATPase and Gfp was functional and oscillated in nucleoid regions on a time scale of minutes. ParA-green fluorescent protein (Gfp) oscillation depended on both ParB and parC but was independent of minCDE. Point mutations in the Walker A box motif simultaneously abolished plasmid stabilization and ParA-Gfp oscillation. These observations raise the possibility that ParA oscillation is prerequisite for active plasmid segregation.
Figures
References
-
- Hiraga S. Annu Rev Genet. 2000;34:21–59. - PubMed
-
- Gordon G S, Wright A. Annu Rev Microbiol. 2000;54:681–708. - PubMed
-
- Møller-Jensen J, Jensen R B, Gerdes K. Trends Microbiol. 2000;8:313–320. - PubMed
-
- Gerdes K, Møller-Jensen J, Bugge J R. Mol Microbiol. 2000;37:455–466. - PubMed
-
- Watanabe E, Wachi M, Yamasaki M, Nagai K. Mol Gen Genet. 1992;234:346–352. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
