

Protein kinase modulation of a neuronal cation channel requires protein-protein interactions mediated by an Src homology 3 domain
- PMID: 11756482
- PMCID: PMC6757624
- DOI: 10.1523/JNEUROSCI.22-01-00001.2002
Protein kinase modulation of a neuronal cation channel requires protein-protein interactions mediated by an Src homology 3 domain
Abstract
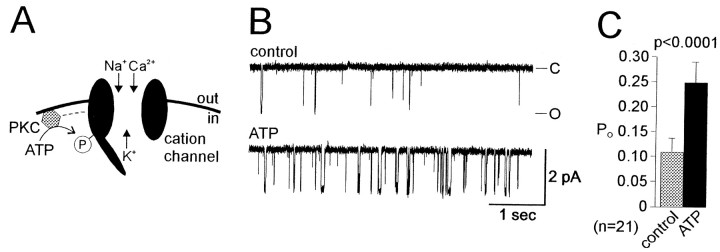
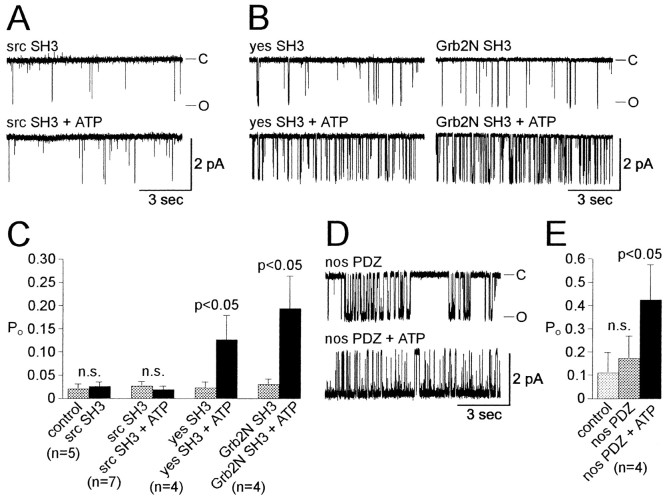
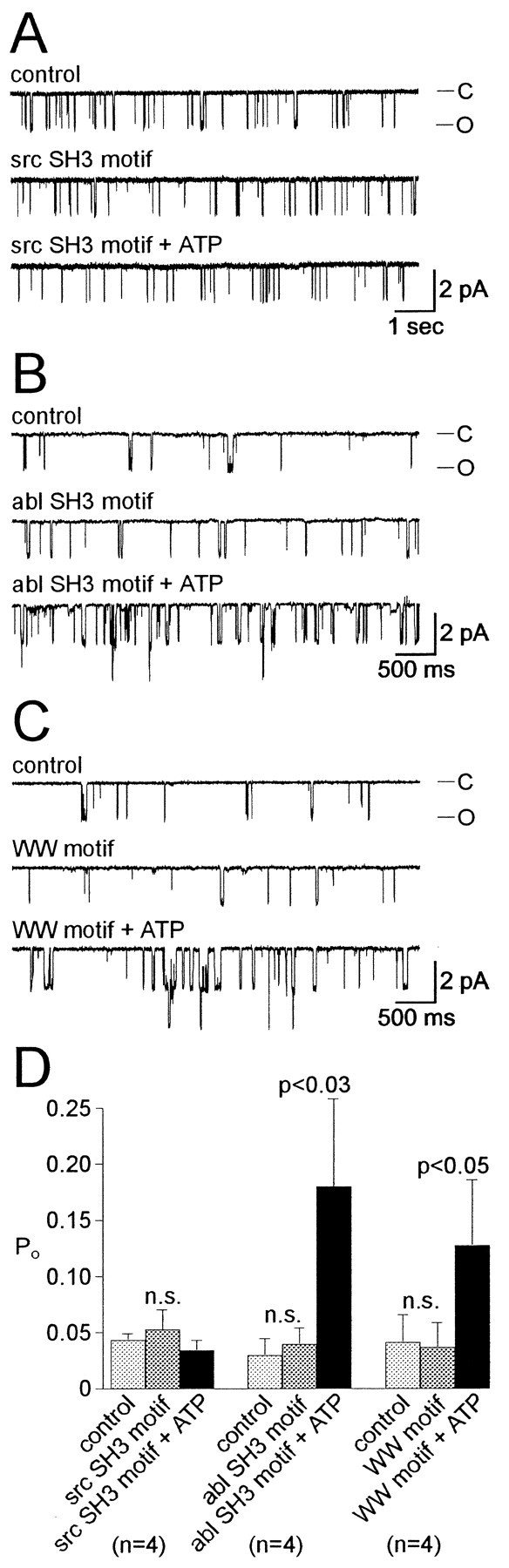
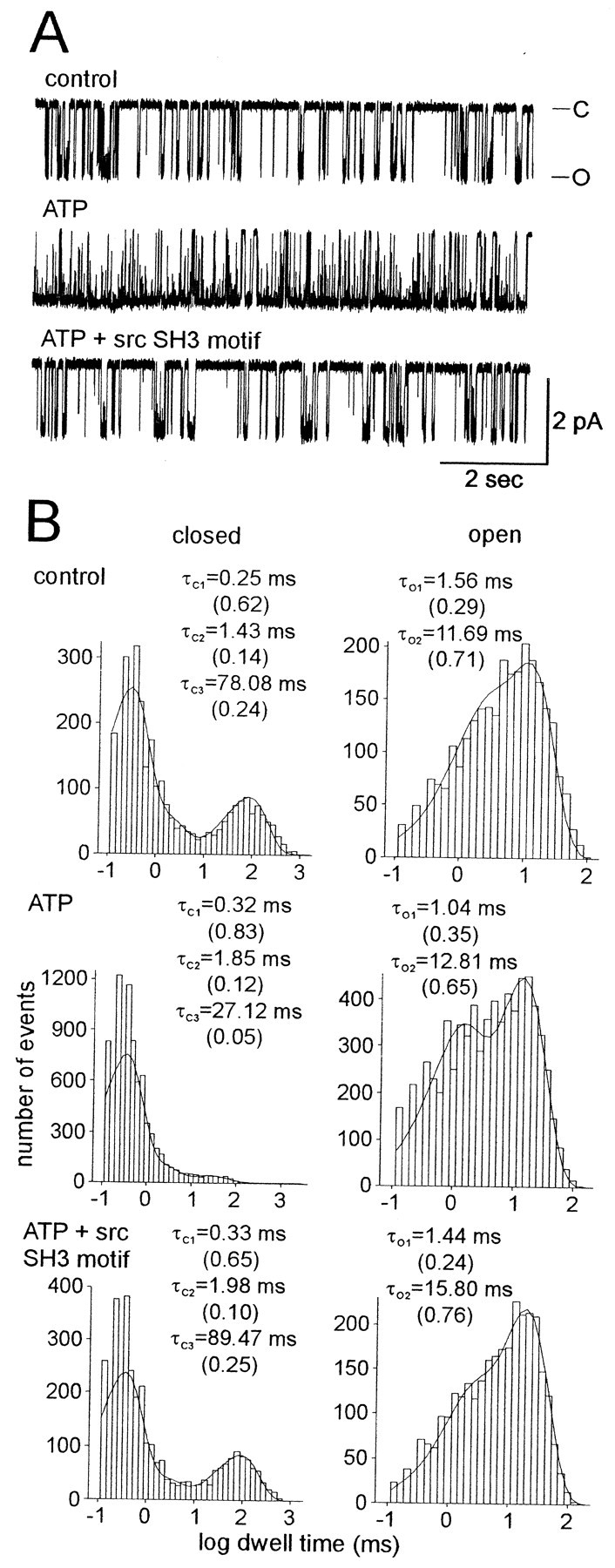
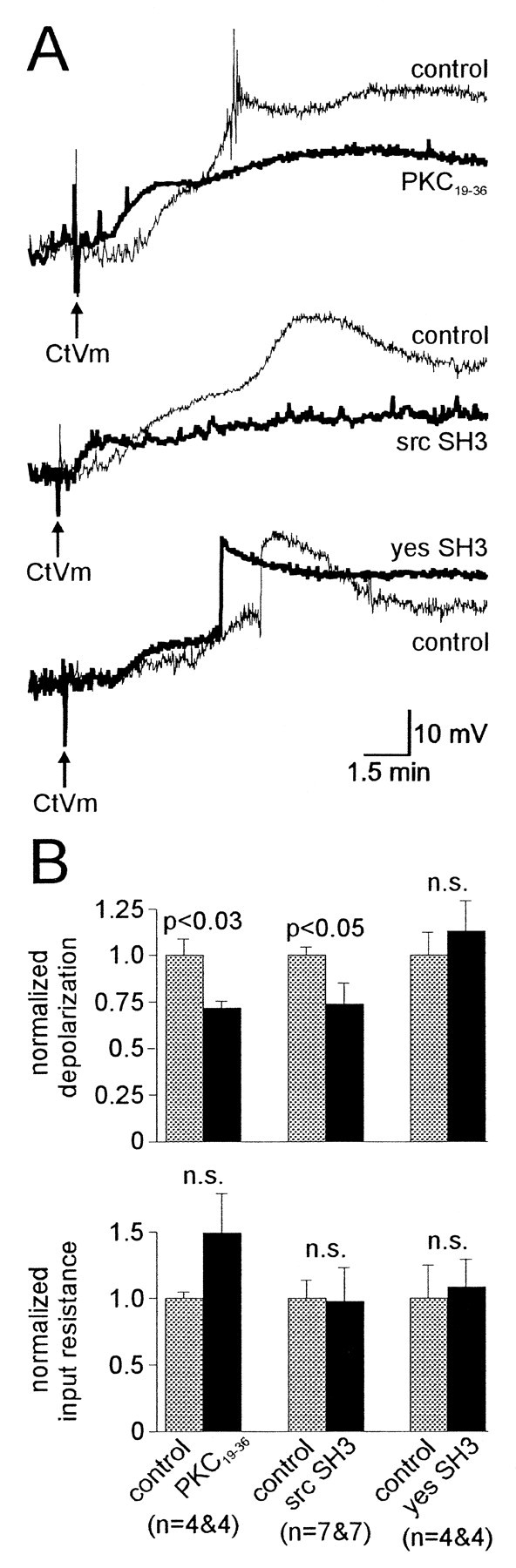
Accumulating evidence suggests that many ion channels reside within a multiprotein complex that contains kinases and other signaling molecules. The role of the adaptor proteins that physically link these complexes together for the purposes of ion channel modulation, however, has been little explored. Here, we examine the protein-protein interactions required for regulation of an Aplysia bag cell neuron cation channel by a closely associated protein kinase C (PKC). In inside-out patches, the PKC-dependent enhancement of cation channel open probability could be prevented by the src homology 3 (SH3) domain, presumably by disrupting a link between the channel and the kinase. SH3 and PDZ domains from other proteins were ineffective. Modulation was also prevented by an SH3 motif peptide that preferentially binds the SH3 domain of src. Furthermore, whole-cell depolarizations elicited by cation channel activation were decreased by the src SH3 domain. These data suggest that the cation channel-PKC association may require SH3 domain-mediated interactions to bring about modulation, promote membrane depolarization, and initiate prolonged changes in bag cell neuron excitability. In general, protein-protein interactions between ion channels and protein kinases may be a prominent mechanism underlying neuromodulation.
Figures
Similar articles
-
Regulation of an Aplysia bag-cell neuron cation channel by closely associated protein kinase A and a protein phosphatase.J Neurosci. 2004 Jul 28;24(30):6833-41. doi: 10.1523/JNEUROSCI.1694-04.2004. J Neurosci. 2004. PMID: 15282289 Free PMC article.
-
Regulation of cation channel voltage and Ca2+ dependence by multiple modulators.J Neurophysiol. 2009 Jul;102(1):259-71. doi: 10.1152/jn.00065.2009. Epub 2009 Apr 22. J Neurophysiol. 2009. PMID: 19386758
-
Association/dissociation of a channel-kinase complex underlies state-dependent modulation.J Neurosci. 2005 Aug 31;25(35):8037-47. doi: 10.1523/JNEUROSCI.1903-05.2005. J Neurosci. 2005. PMID: 16135761 Free PMC article.
-
The 5-HT3 receptor channel.Annu Rev Physiol. 1995;57:447-68. doi: 10.1146/annurev.ph.57.030195.002311. Annu Rev Physiol. 1995. PMID: 7539990 Review. No abstract available.
-
Signalling synapse formation between identified neurons.J Physiol Paris. 1995;89(3):115-23. doi: 10.1016/0928-4257(96)80108-2. J Physiol Paris. 1995. PMID: 7581300 Review.
Cited by
-
Prolonged activation of Ca2+-activated K+ current contributes to the long-lasting refractory period of Aplysia bag cell neurons.J Neurosci. 2002 Dec 1;22(23):10134-41. doi: 10.1523/JNEUROSCI.22-23-10134.2002. J Neurosci. 2002. PMID: 12451114 Free PMC article.
-
Regulation of recombinant and native hyperpolarization-activated cation channels.Mol Neurobiol. 2004 Dec;30(3):279-305. doi: 10.1385/MN:30:3:279. Mol Neurobiol. 2004. PMID: 15655253 Review.
-
Ca2+-dependent regulation of a non-selective cation channel from Aplysia bag cell neurones.J Physiol. 2006 Sep 1;575(Pt 2):491-506. doi: 10.1113/jphysiol.2006.105833. Epub 2006 Jun 8. J Physiol. 2006. PMID: 16763004 Free PMC article.
-
Regulation of neuronal excitability by interaction of fragile X mental retardation protein with slack potassium channels.J Neurosci. 2012 Oct 31;32(44):15318-27. doi: 10.1523/JNEUROSCI.2162-12.2012. J Neurosci. 2012. PMID: 23115170 Free PMC article.
-
Identification of a functional interaction between Kv4.3 channels and c-Src tyrosine kinase.Biochim Biophys Acta. 2008 Oct;1783(10):1884-92. doi: 10.1016/j.bbamcr.2008.06.011. Epub 2008 Jun 20. Biochim Biophys Acta. 2008. PMID: 18620005 Free PMC article.
References
-
- Alonso A, Llinas RR. Subthreshold Na+-dependent theta-like rhythmicity in stellate cells of entorhinal cortex layer II. Nature. 1989;342:175–177. - PubMed
-
- Chung S, Reinhart PH, Martin BL, Brautigan D, Levitan IB. Protein kinase activity closely associated with a reconstituted calcium-activated potassium channel. Science. 1991;253:560–562. - PubMed
-
- Coghlan VM, Perrino BA, Howard M, Langeberg LK, Hicks JB, Gallatin WM, Scott JD. Association of protein kinase A and protein phosphatase 2B with a common anchoring protein. Science. 1995;267:108–111. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous