

Assessing the role of calcium-induced calcium release in short-term presynaptic plasticity at excitatory central synapses
- PMID: 11756484
- PMCID: PMC6757598
- DOI: 10.1523/JNEUROSCI.22-01-00021.2002
Assessing the role of calcium-induced calcium release in short-term presynaptic plasticity at excitatory central synapses
Abstract
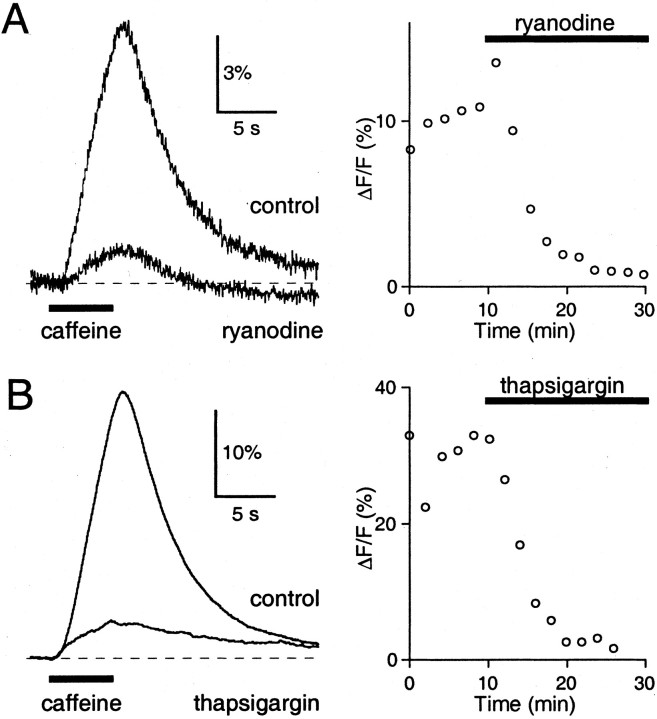
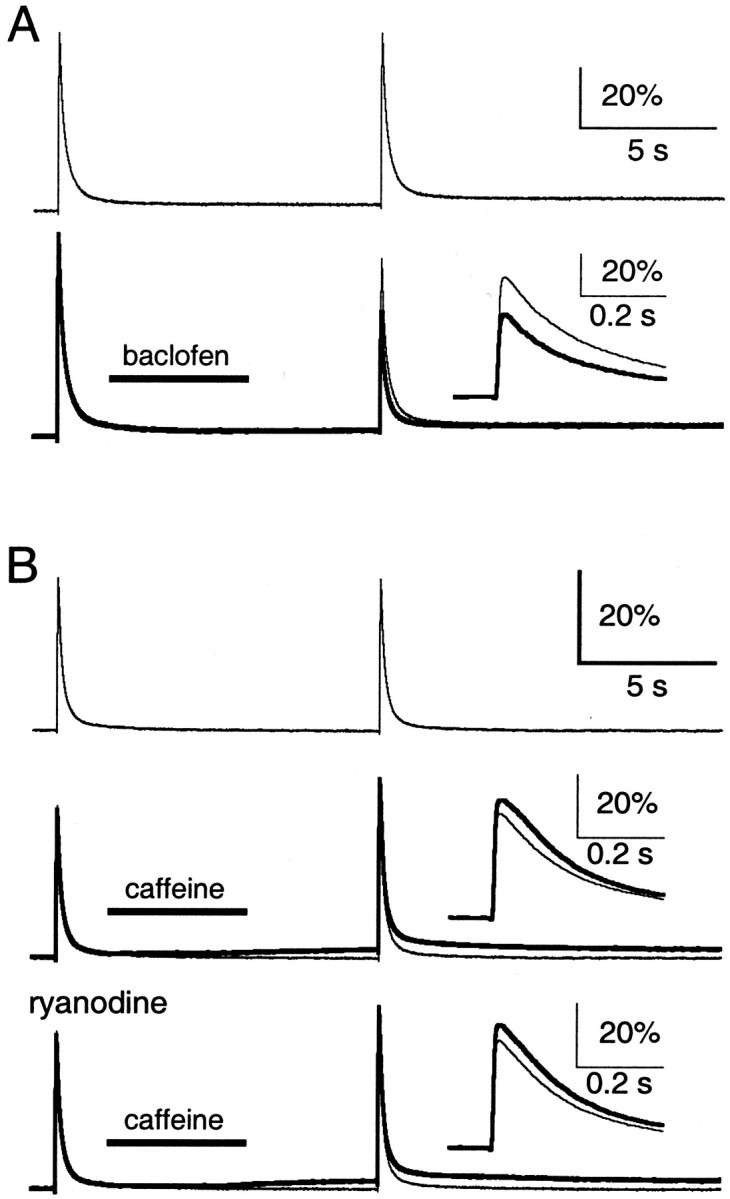
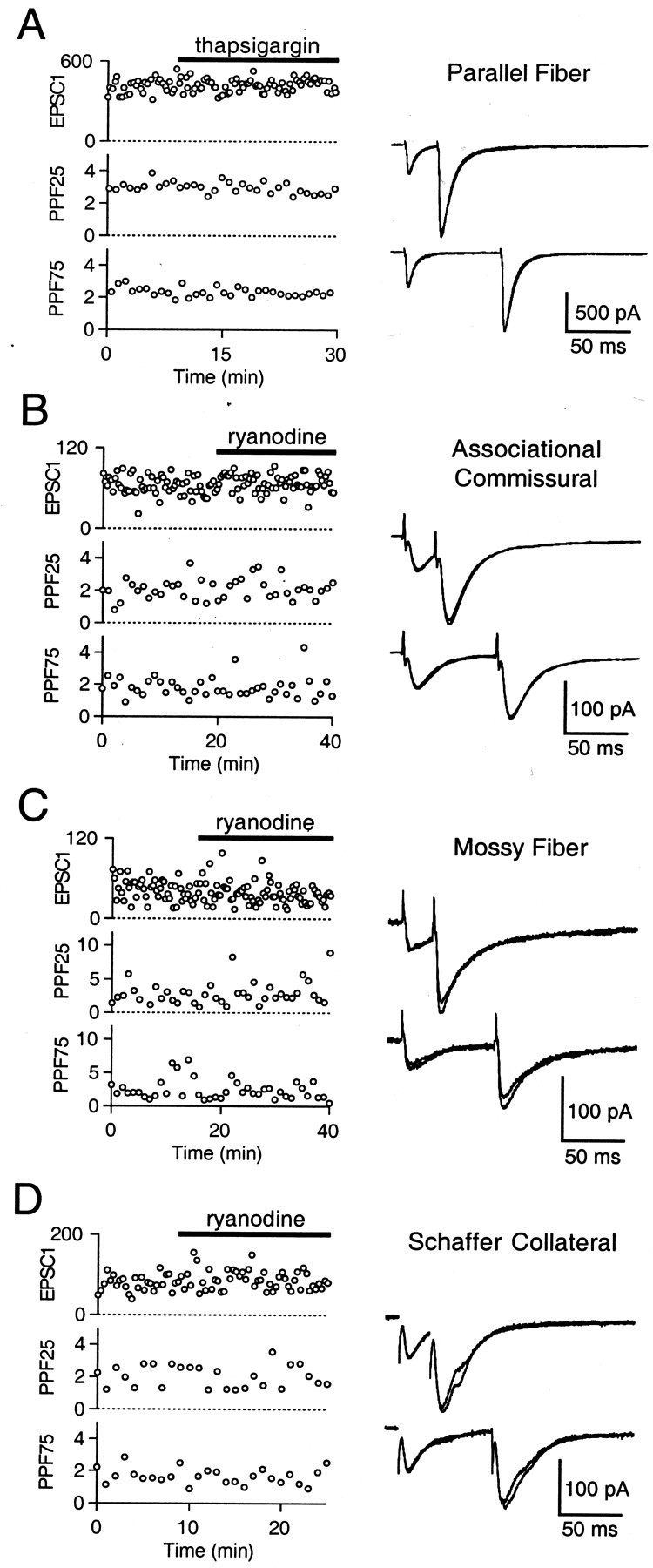
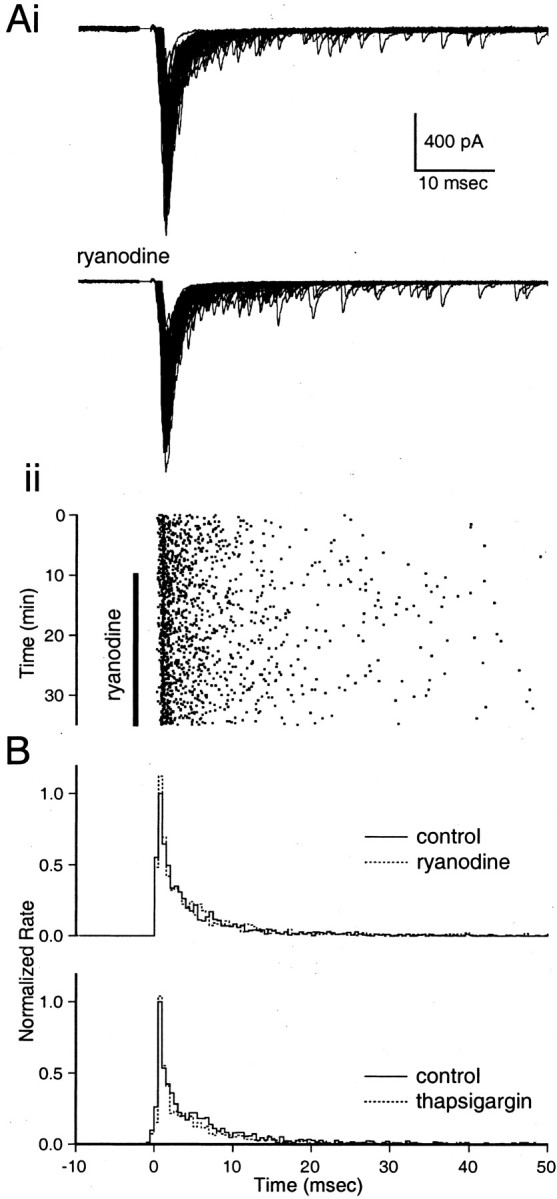
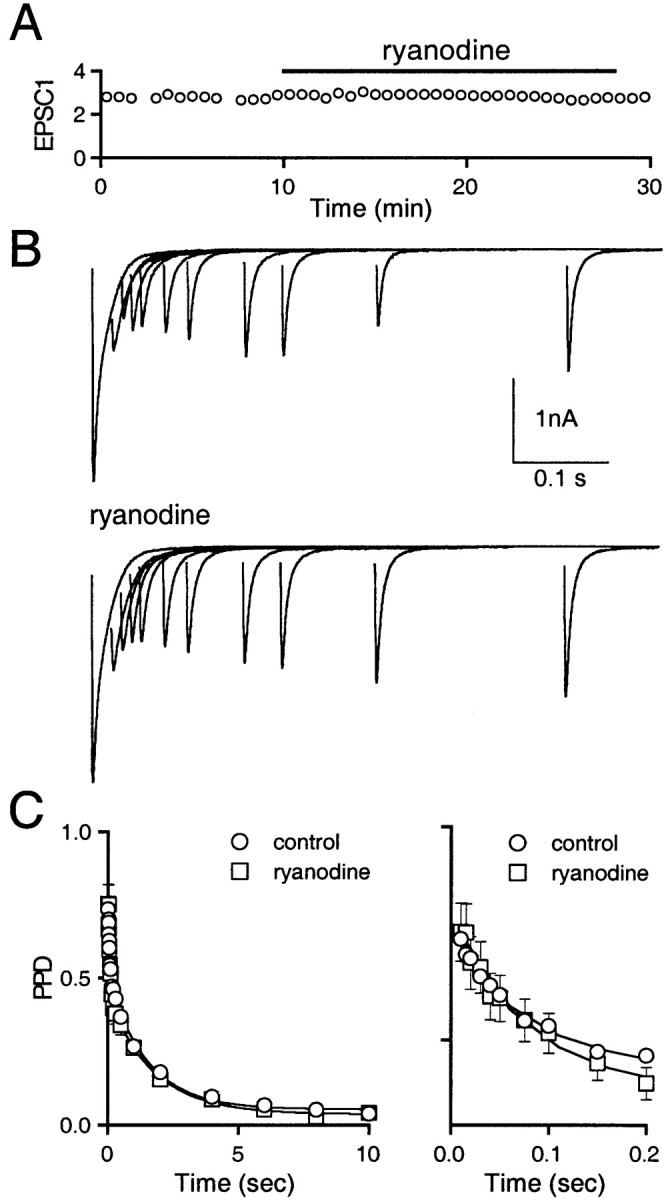
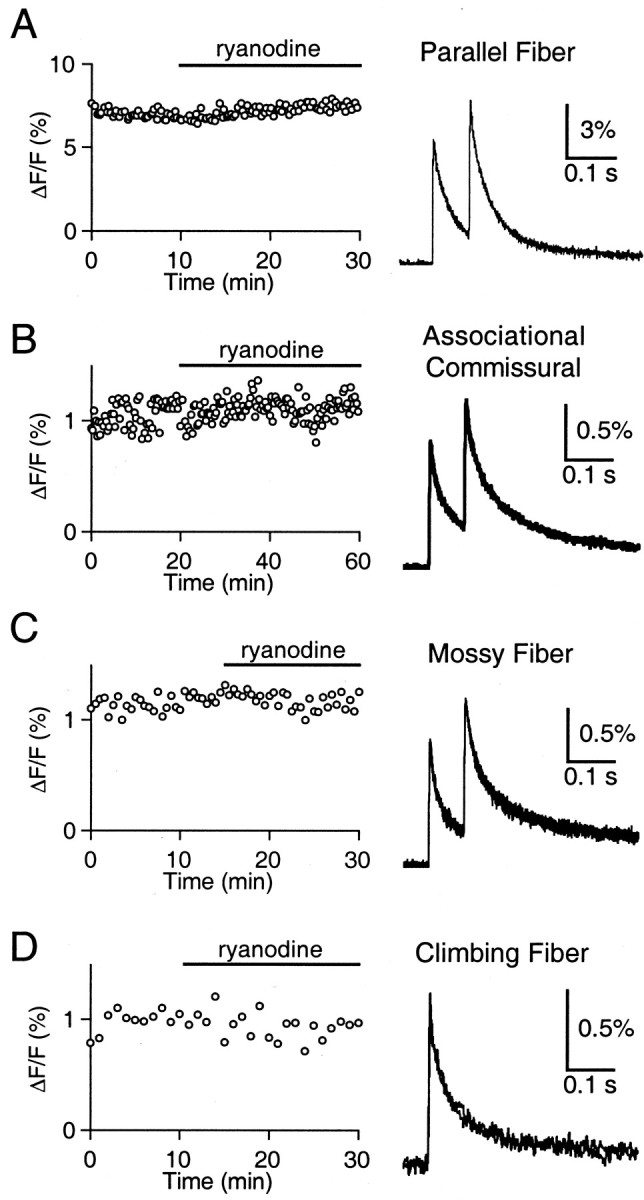
Recent evidence suggests that internal calcium stores and calcium-induced calcium release (CICR) provide an important source of calcium that drives short-term presynaptic plasticity at central synapses. Here we tested for the involvement of CICR in short-term presynaptic plasticity at six excitatory synapses in acute rat hippocampal and cerebellar brain slices. Depletion of internal calcium stores with thapsigargin and prevention of CICR with ryanodine have no effect on paired-pulse facilitation, delayed release of neurotransmitter, or calcium-dependent recovery from depression. Fluorometric calcium measurements also show that these drugs have no effect on the residual calcium signal that underlies these forms of short-term presynaptic plasticity. Finally, although caffeine causes CICR in Purkinje cell bodies and dendrites, it does not elicit CICR in parallel fiber inputs to these cells. Taken together, these results indicate that for the excitatory synapses studied here, internal calcium stores and CICR do not contribute to short-term presynaptic plasticity on the milliseconds-to-seconds time scale. Instead, this plasticity is driven by the residual calcium signal arising from calcium entry through voltage-gated calcium channels.
Figures
References
-
- Araque A, Carmignoto G, Haydon PG. Dynamic signaling between astrocytes and neurons. Annu Rev Physiol. 2001;63:795–813. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources