

Replication initiation patterns in the beta-globin loci of totipotent and differentiated murine cells: evidence for multiple initiation regions
- PMID: 11756541
- PMCID: PMC139749
- DOI: 10.1128/MCB.22.2.442-452.2002
Replication initiation patterns in the beta-globin loci of totipotent and differentiated murine cells: evidence for multiple initiation regions
Abstract
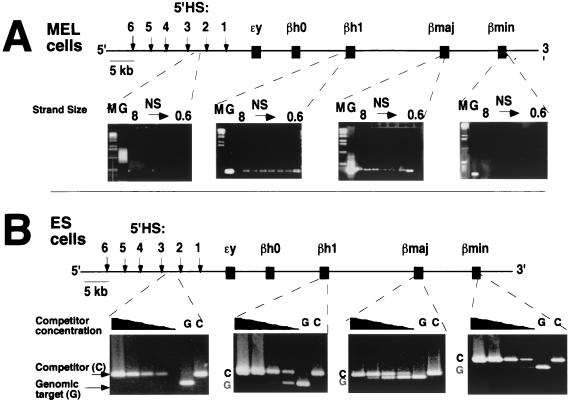
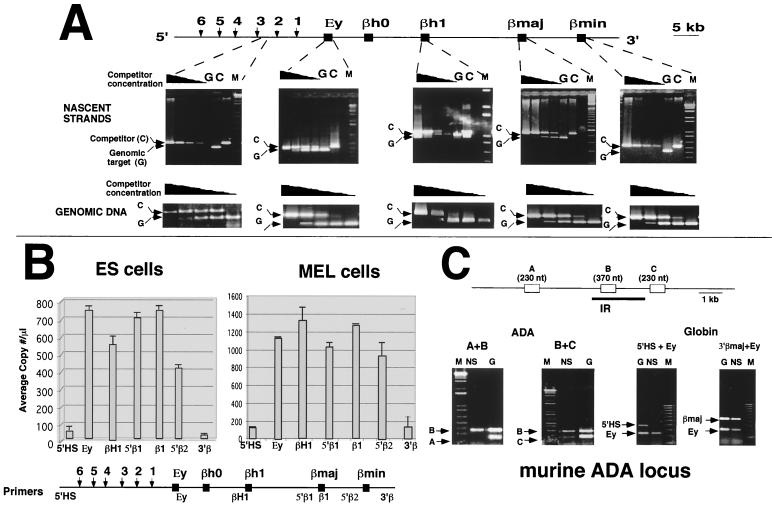
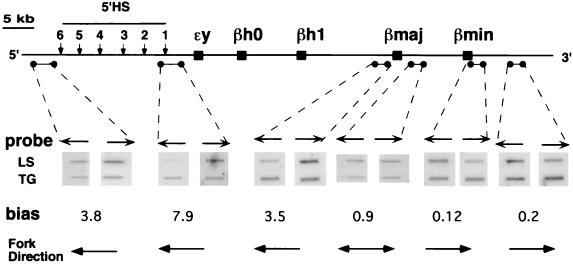
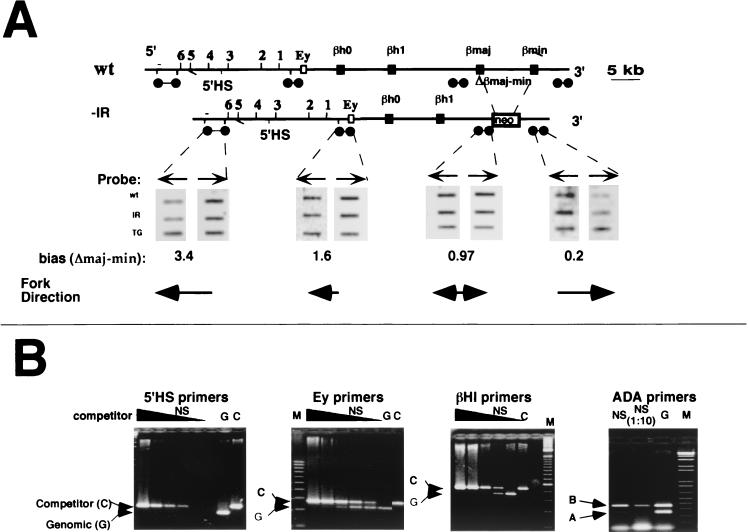
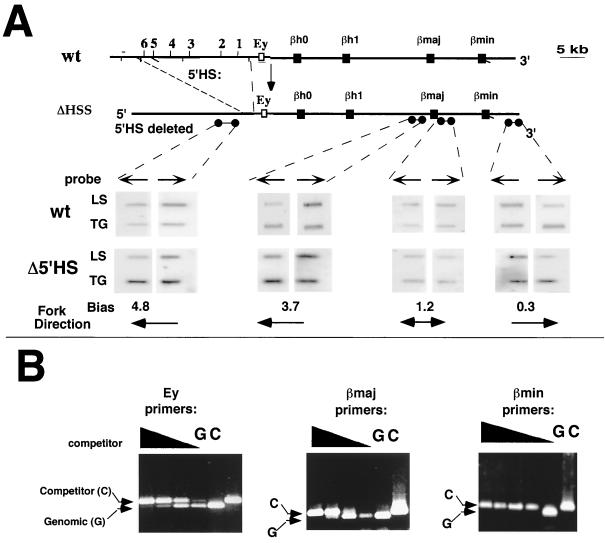
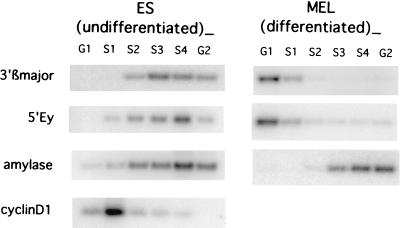
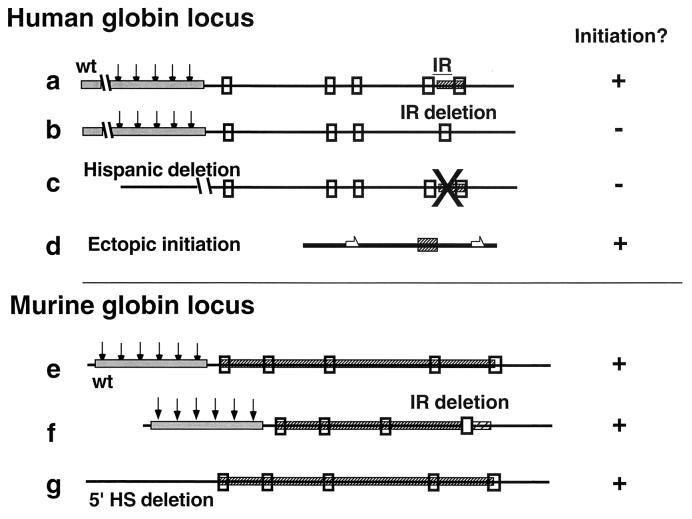
The replication initiation pattern of the murine beta-globin locus was analyzed in totipotent embryonic stem cells and in differentiated cell lines. Initiation events in the murine beta-globin locus were detected in a region extending from the embryonic Ey gene to the adult betaminor gene, unlike the restricted initiation observed in the human locus. Totipotent and differentiated cells exhibited similar initiation patterns. Deletion of the region between the adult globin genes did not prevent initiation in the remainder of the locus, suggesting that the potential to initiate DNA replication was not contained exclusively within the primary sequence of the deleted region. In addition, a deletion encompassing the six identified 5' hypersensitive sites in the mouse locus control region had no effect on initiation from within the locus. As this deletion also did not affect the chromatin structure of the locus, we propose that the sequences determining both chromatin structure and replication initiation lie outside the hypersensitive sites removed by the deletion.
Figures
References
-
- Aladjem, M., L.-W. Rodewald, J. L. Kolman, and G. M. Wahl. 1998. Genetic dissection of a mammalian replicator in the human beta-globin locus. Science 281:1005–1009. - PubMed
-
- Aladjem, M., and G. M. Wahl. 1997. Mapping replication origins by leading strand analysis in the absence of protein synthesis. Methods Companion Methods Enzymol. 13:281–292. - PubMed
-
- Aladjem, M. I., M. Groudine, L. L. Brody, E. S. Dieken, R. E. K. Fournier, G. M. Wahl, and E. M. Epner. 1995. Participation of the human beta globin locus control region in initiation of DNA replication. Science 270:815–819. - PubMed
-
- Aladjem, M. I., B. T. Spike, L. W. Rodewald, T. J. Hope, M. Klemm, R. Jaenisch, and G. M. Wahl. 1998. ES cells do not activate p53-dependent stress responses and undergo p53-independent apoptosis in response to DNA damage. Curr. Biol. 8:145–155. - PubMed
-
- Bielinsky, A. K., and S. A. Gerbi. 1998. Discrete start sites for DNA synthesis in the yeast ARS1 origin. Science 279:95–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials