

Single-base-pair discrimination of terminal mismatches by using oligonucleotide microarrays and neural network analyses
- PMID: 11772632
- PMCID: PMC126557
- DOI: 10.1128/AEM.68.1.235-244.2002
Single-base-pair discrimination of terminal mismatches by using oligonucleotide microarrays and neural network analyses
Abstract
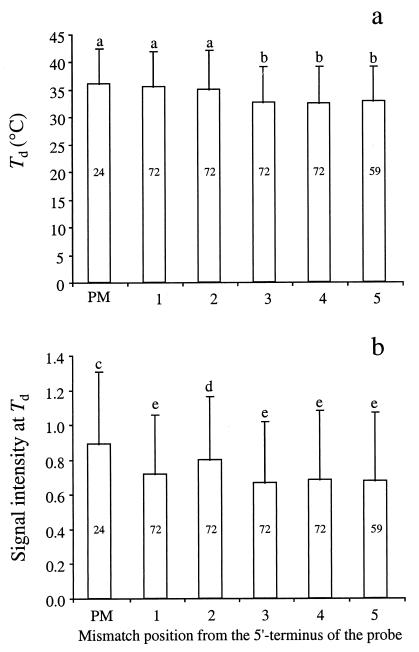
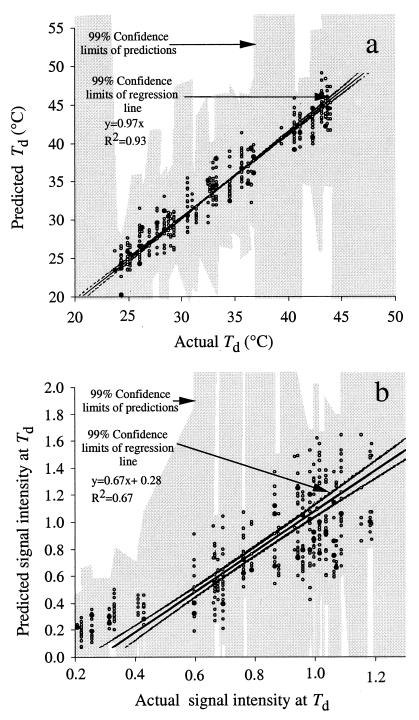
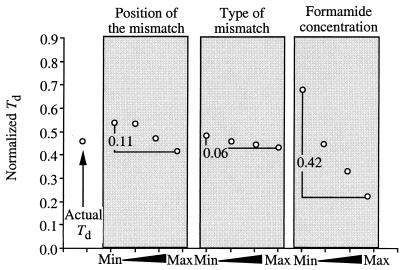
The effects of single-base-pair near-terminal and terminal mismatches on the dissociation temperature (T(d)) and signal intensity of short DNA duplexes were determined by using oligonucleotide microarrays and neural network (NN) analyses. Two perfect-match probes and 29 probes having a single-base-pair mismatch at positions 1 to 5 from the 5' terminus of the probe were designed to target one of two short sequences representing 16S rRNA. Nonequilibrium dissociation rates (i.e., melting profiles) of all probe-target duplexes were determined simultaneously. Analysis of variance revealed that position of the mismatch, type of mismatch, and formamide concentration significantly affected the T(d) and signal intensity. Increasing the concentration of formamide in the washing buffer decreased the T(d) and signal intensity, and it decreased the variability of the signal. Although T(d)s of probe-target duplexes with mismatches in the first or second position were not significantly different from one another, duplexes with mismatches in the third to fifth positions had significantly lower T(d)s than those with mismatches in the first or second position. The trained NNs predicted the T(d) with high accuracies (R(2) = 0.93). However, the NNs predicted the signal intensity only moderately accurately (R(2) = 0.67), presumably due to increased noise in the signal intensity at low formamide concentrations. Sensitivity analysis revealed that the concentration of formamide explained most (75%) of the variability in T(d)s, followed by position of the mismatch (19%) and type of mismatch (6%). The results suggest that position of the mismatch at or near the 5' terminus plays a greater role in determining the T(d) and signal intensity of duplexes than the type of mismatch.
Figures
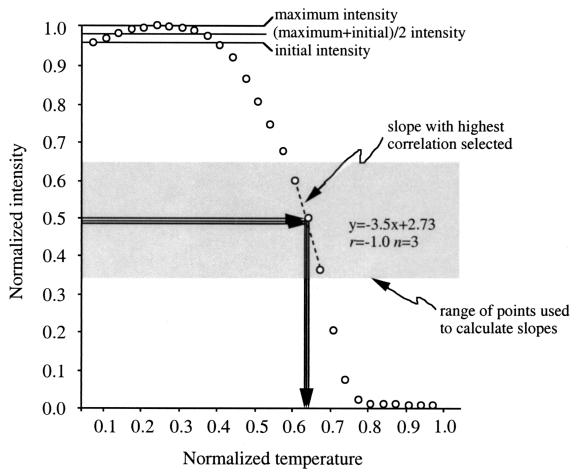
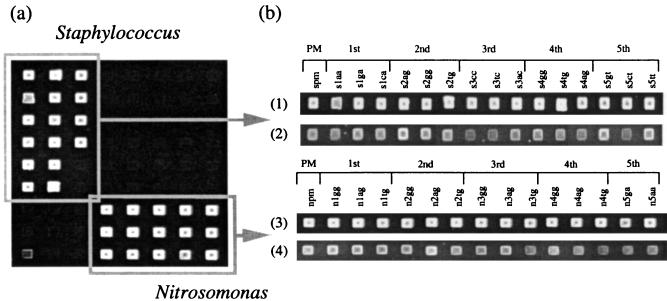
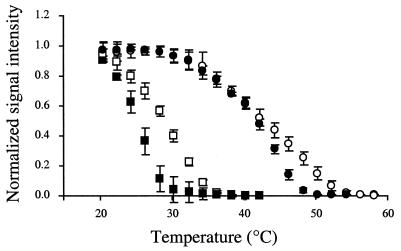
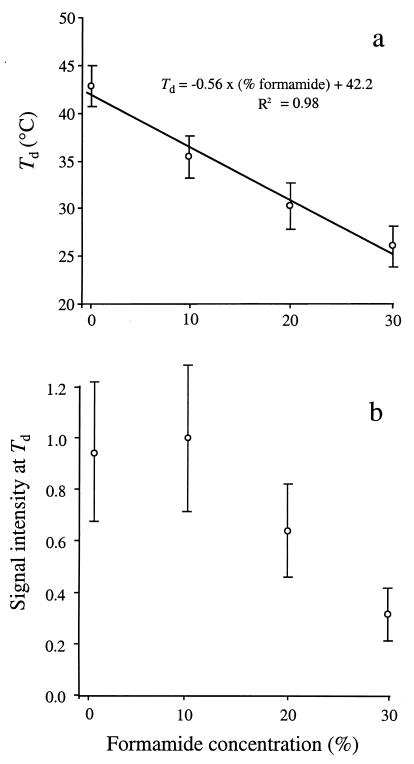
References
-
- Aleksander, I., and H. Morton. 1991. An introduction to neural computing, p.1–20. Chapman & Hall, Ltd., London, United Kingdom.
-
- Almeida, J. S., and P. A. Noble. 2000. Neural computing in microbiology. J. Microbiol. Methods 43:1–2. - PubMed
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215:403–410. - PubMed
-
- Basheer, I. A., and M. Hajmeer. 2000. Artificial neural networks: fundamentals, computing, design, and application. J. Microbiol. Methods 43:3–31. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
