

Artificial extension of the adenovirus fiber shaft inhibits infectivity in coxsackievirus and adenovirus receptor-positive cell lines
- PMID: 11773386
- PMCID: PMC135866
- DOI: 10.1128/jvi.76.3.1100-1108.2002
Artificial extension of the adenovirus fiber shaft inhibits infectivity in coxsackievirus and adenovirus receptor-positive cell lines
Abstract
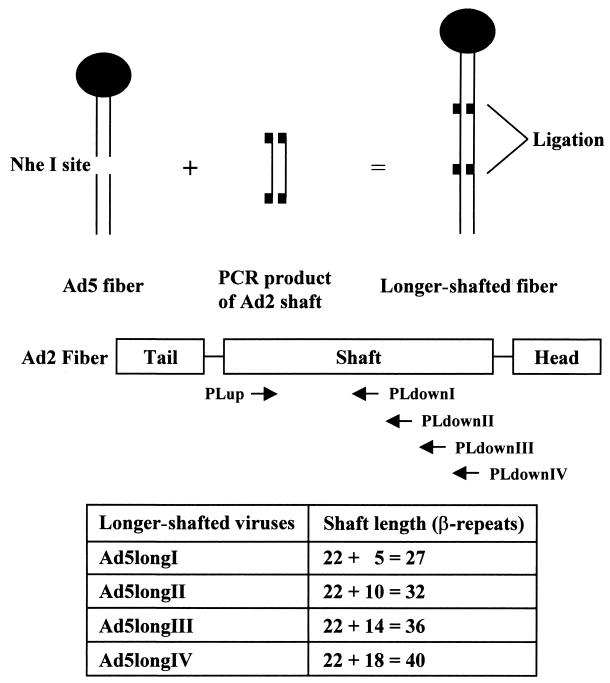
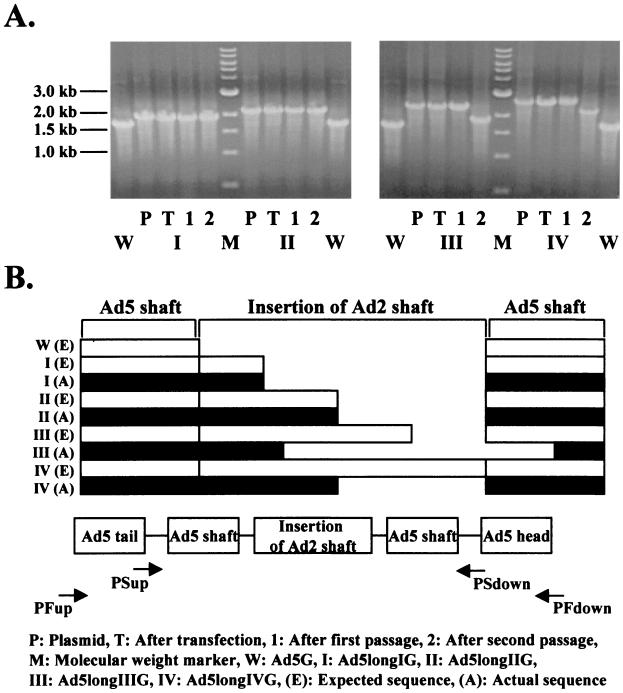
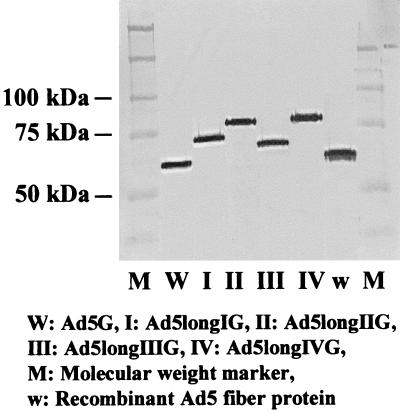
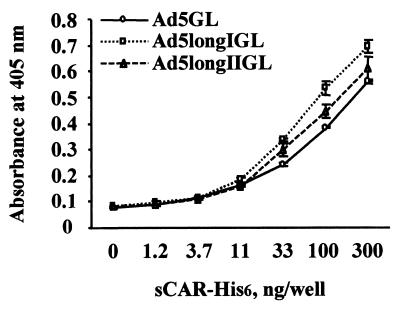
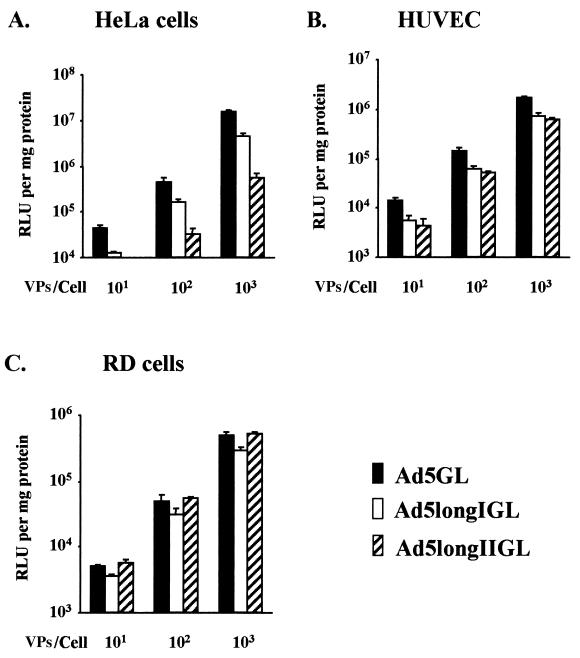
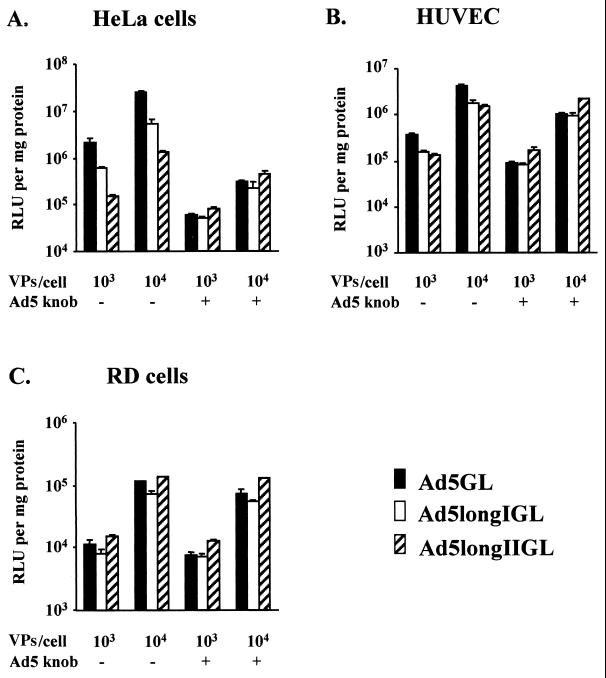
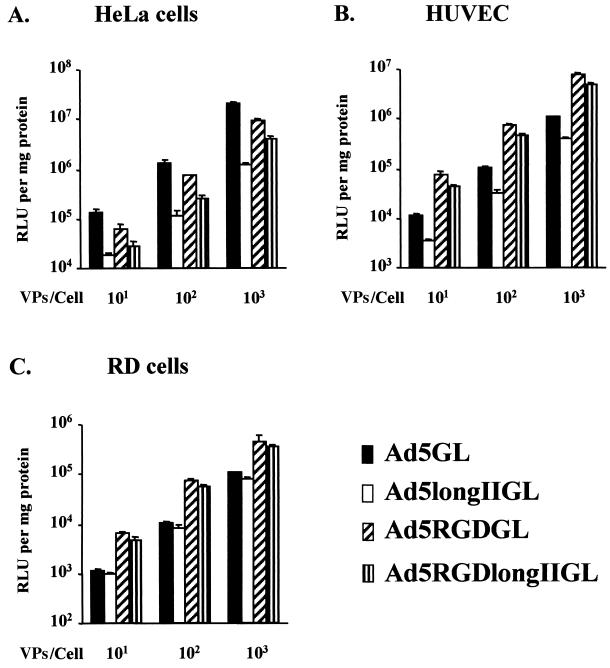
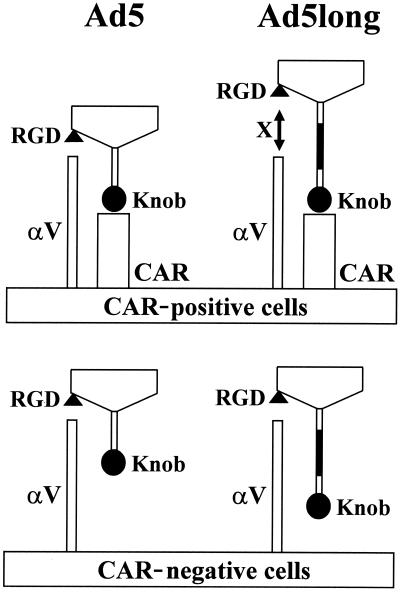
Recent studies demonstrate that virus-cellular receptor interactions are not the sole determinants of adenovirus (Ad) tropism. It has been shown that the fiber shaft length, which ranges from 6 to 23 beta-repeats in human Ads, also influences viral tropism. However, there is no report that investigates whether artificial extension of the shaft alters the infectivity profile of Ad. Therefore, we constructed Ad serotype 5 (Ad5) capsid-based longer-shafted Ad vectors by incorporating Ad2 shaft fragments of different lengths into the Ad5 shaft. We show that "longer-shafted" Ad vectors (up to 32 beta-repeats) could be rescued. We also show that longer-shafted Ad vectors had no impact on knob-CAR (coxsackievirus and Ad receptor) interaction compared to wild-type Ad. Nevertheless, gene transfer efficiencies of longer-shafted Ad vectors were lower in CAR-positive cell lines compared to wild-type Ad. We suggest that artificial extension of the shaft can inhibit infectivity in the context of CAR-positive cell lines without modification of knob-CAR interaction.
Figures
References
-
- Chroboczek, J., R. W. H. Ruigrok, and S. Cusack. 1995. Adenovirus fiber. Curr. Top. Microbiol. Immunol. 199:163–200. - PubMed
-
- Crompton, J., C. I. Toogood, N. Wallis, and R. T. Hay. 1994. Expression of a foreign epitope on the surface of the adenovirus hexon. J. Gen. Virol. 75:133–139. - PubMed
-
- Dmitriev, I., V. Krasnykh, C. R. Miller, M. Wang, E. Kashentseva, G. Mikheeva, N. Belousova, and D. T. Curiel. 1998. An adenovirus vector with genetically modified fibers demonstrates expanded tropism via utilization of a coxsackievirus and adenovirus receptor-independent cell entry mechanism. J. Virol. 72:9706–9713. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
