

Fyn tyrosine kinase is a downstream mediator of Rho/PRK2 function in keratinocyte cell-cell adhesion
- PMID: 11777936
- PMCID: PMC2173591
- DOI: 10.1083/jcb.200105140
Fyn tyrosine kinase is a downstream mediator of Rho/PRK2 function in keratinocyte cell-cell adhesion
Abstract
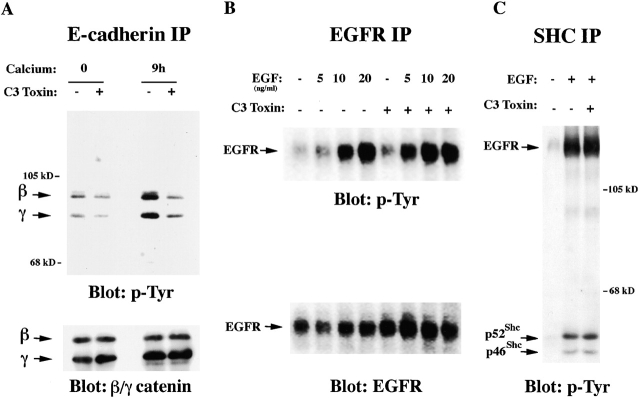
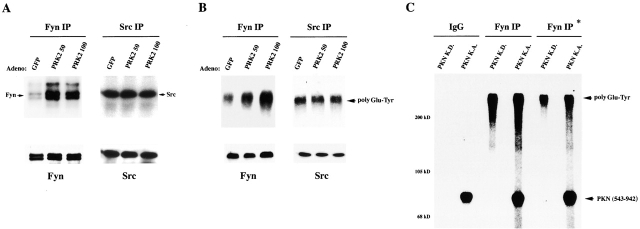
The Rho GTPase and Fyn tyrosine kinase have been implicated previously in positive control of keratinocyte cell-cell adhesion. Here, we show that Rho and Fyn operate along the same signaling pathway. Endogenous Rho activity increases in differentiating keratinocytes and is required for both Fyn kinase activation and increased tyrosine phosphorylation of beta- and gamma-catenin, which is associated with the establishment of keratinocyte cell-cell adhesion. Conversely, expression of constitutive active Rho is sufficient to promote cell-cell adhesion through a tyrosine kinase- and Fyn-dependent mechanism, trigger Fyn kinase activation, and induce tyrosine phosphorylation of beta- and gamma-catenin and p120ctn. The positive effects of activated Rho on cell-cell adhesion are not induced by an activated Rho mutant with defective binding to the serine/threonine PRK2/PKN kinases. Endogenous PRK2 kinase activity increases with keratinocyte differentiation, and, like activated Rho, increased PRK2 activity promotes keratinocyte cell-cell adhesion and induces tyrosine phosphorylation of beta- and gamma-catenin and Fyn kinase activation. Thus, these findings reveal a novel role of Fyn as a downstream mediator of Rho in control of keratinocyte cell-cell adhesion and implicate the PRK2 kinase, a direct Rho effector, as a link between Rho and Fyn activation.
Figures













References
-
- Anastasiadis, P.Z., S.Y. Moon, M.A. Thoreson, D.J. Mariner, H.C. Crawford, Y. Zheng, and A.B. Reynolds. 2000. Inhibition of RhoA by p120 catenin. Nat. Cell Biol. 2:637–644. - PubMed
-
- Anastasiadis, P.Z., and A.B. Reynolds. 2000. The p120 catenin family: complex roles in adhesion, signaling and cancer. J. Cell Sci. 113:1319–1334. - PubMed
-
- Behrens, J., L. Vakaet, R. Friis, E. Winterhager, F. Van Roy, M.M. Mareel, and W. Birchmeier. 1993. Loss of epithelial differentiation and gain of invasiveness correlates with tyrosine phosphorylation of the E-cadherin/beta-catenin complex in cells transformed with a temperature-sensitive v-SRC gene. J. Cell Biol. 120:757–766. - PMC - PubMed
-
- Belsches, A.P., M.D. Haskell, and S.J. Parsons. 1997. Role of c-Src tyrosine kinase in EGF-induced mitogenesis. Front Biosci. 2:501–518. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
