

Antitumor monoclonal antibodies enhance cross-presentation ofcCellular antigens and the generation of myeloma-specific killer T cells by dendritic cells
- PMID: 11781371
- PMCID: PMC2196013
- DOI: 10.1084/jem.20011097
Antitumor monoclonal antibodies enhance cross-presentation ofcCellular antigens and the generation of myeloma-specific killer T cells by dendritic cells
Abstract
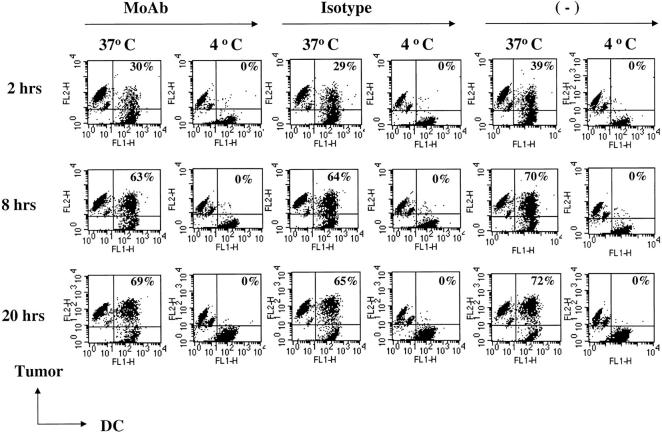
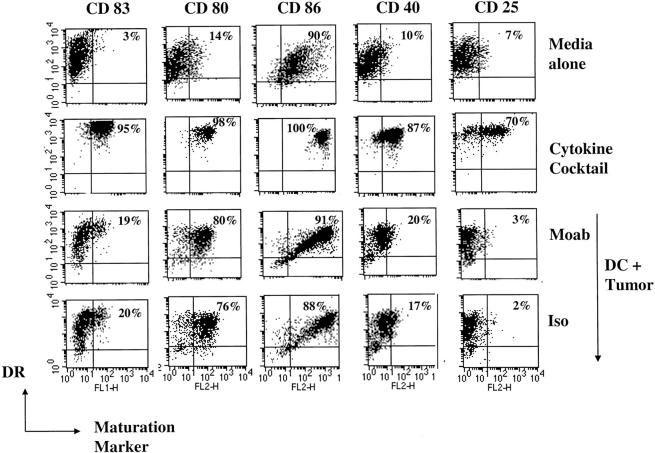
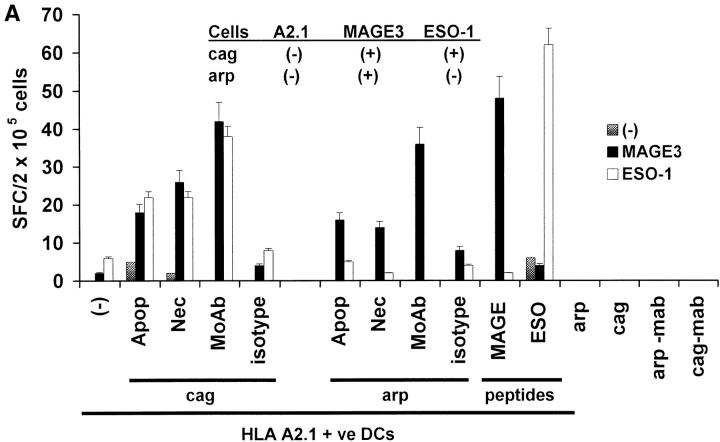
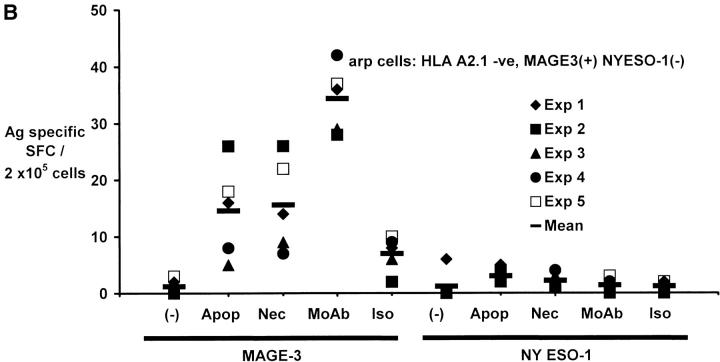
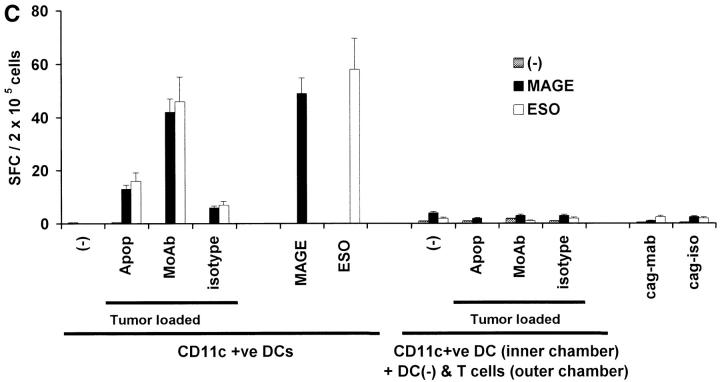
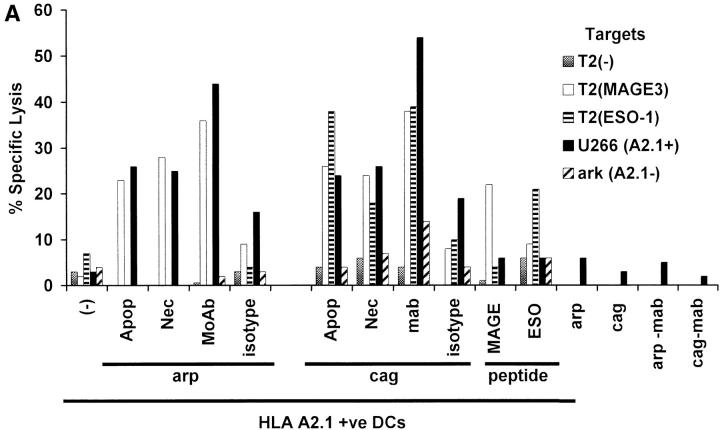
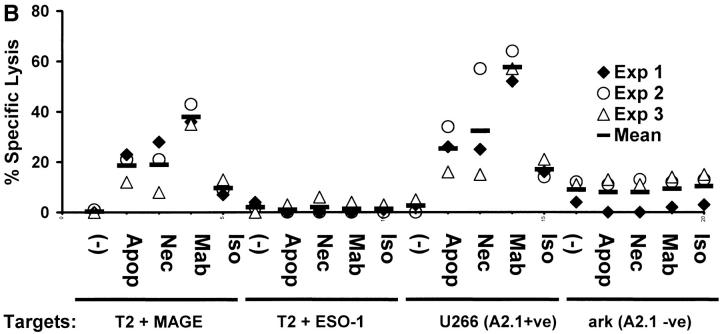
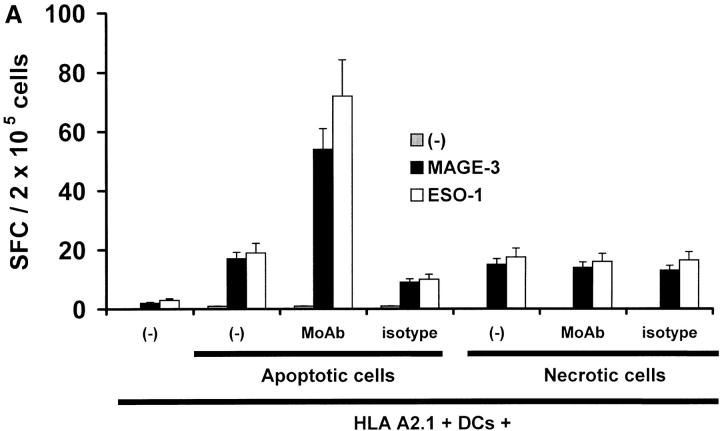
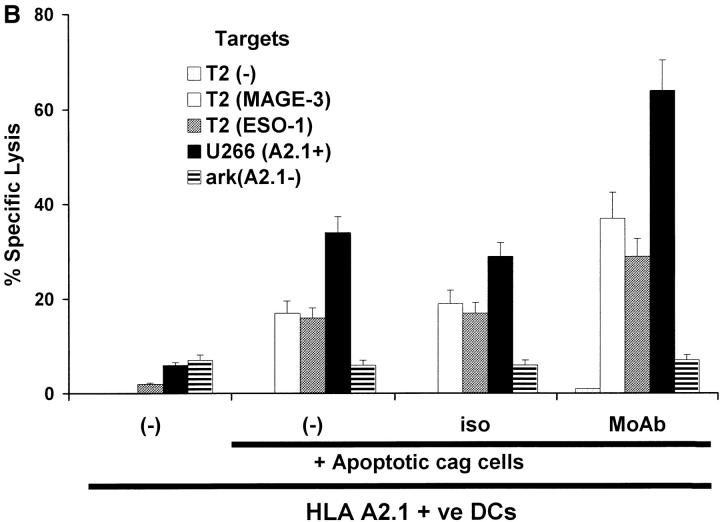
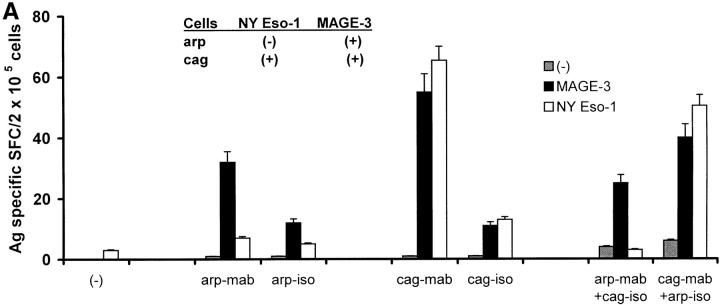
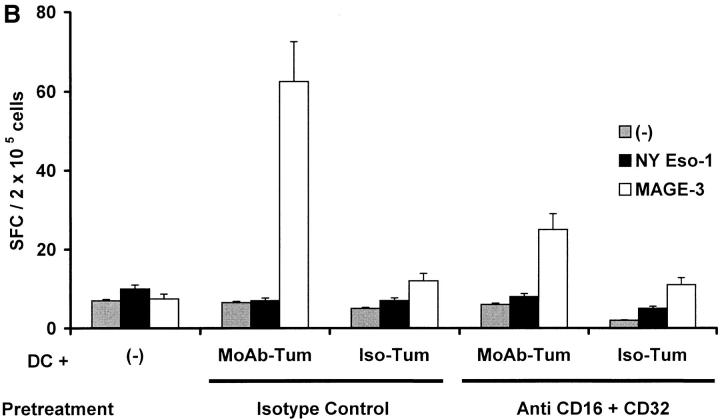
The mechanism of antitumor effect of monoclonal antibodies (mAbs) is not fully understood. Here we show that coating myeloma cells with anti-syndecan-1 antibody promotes cross-presentation of cellular antigens by dendritic cells (DCs) to autologous T cells from healthy donors. The tumor cells treated with anti-syndecan-1 or isotype-matched control antibody were fed to HLA-mismatched monocyte-derived immature DCs. Tumor cell-loaded mature DCs induced a strong CD8(+) T cell response that was specific for the cancer-testis (C-T) antigens expressed in the tumor. The CD8(+) T cells killed peptide-pulsed targets, as well as myeloma tumor cells. Importantly, mAbs-coated tumor-loaded DCs were consistently superior to DCs loaded with peptides or dying cells for eliciting tumor-specific killer T cells. This enhanced cross-presentation was not due to enhanced tumor cell uptake or to DC maturation. When mixtures of NY-Eso-1-positive and -negative myeloma cells were captured by DCs, the anti-syndecan-1 antibody had to be on the NY-Eso-1-positive cells to elicit NY-Eso-1-specific response. Cross-presentation was inhibited by pretreatment of DCs with Fc gamma receptor blocking antibodies. Targeting of mAb-coated tumors to DCs may contribute to the efficacy of tumor-reactive mAb and offers a new strategy for immunotherapy.
Figures
Comment in
-
Fc gamma receptors and cross-presentation in dendritic cells.J Exp Med. 2002 Jan 7;195(1):F1-3. doi: 10.1084/jem.20011925. J Exp Med. 2002. PMID: 11781374 Free PMC article. No abstract available.
References
-
- Schubert, U., L.C. Anton, J. Gibbs, C.C. Norbury, J.W. Yewdell, and J.R. Bennink. 2000. Rapid degradation of a large fraction of newly synthesized proteins by proteasomes. Nature. 404:770–774. - PubMed
-
- Heath, W.R., and F.R. Carbone. 2001. Cross-presentation, dendritic cells, tolerance and immunity. Annu. Rev. Immunol. 19:47–64. - PubMed
-
- Berard, F., P. Blanco, J. Davoust, E.M. Neidhart-Berard, M. Nouri-Shirazi, N. Taquet, D. Rimoldi, J.C. Cerottini, J. Banchereau, and A.K. Palucka. 2000. Cross-priming of naive CD8 T cells against melanoma antigens using dendritic cells loaded with killed allogeneic melanoma cells. J. Exp. Med. 192:1535–1544. - PMC - PubMed
-
- Nouri-Shirazi, M., J. Banchereau, D. Bell, S. Burkeholder, E.T. Kraus, J. Davoust, and K.A. Palucka. 2000. Dendritic cells capture killed tumor cells and present their antigens to elicit tumor-specific immune responses. J. Immunol. 165:3797–3803. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
