

Shy1p is necessary for full expression of mitochondrial COX1 in the yeast model of Leigh's syndrome
- PMID: 11782424
- PMCID: PMC125806
- DOI: 10.1093/emboj/21.1.43
Shy1p is necessary for full expression of mitochondrial COX1 in the yeast model of Leigh's syndrome
Abstract
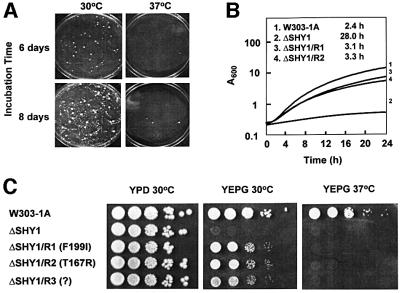
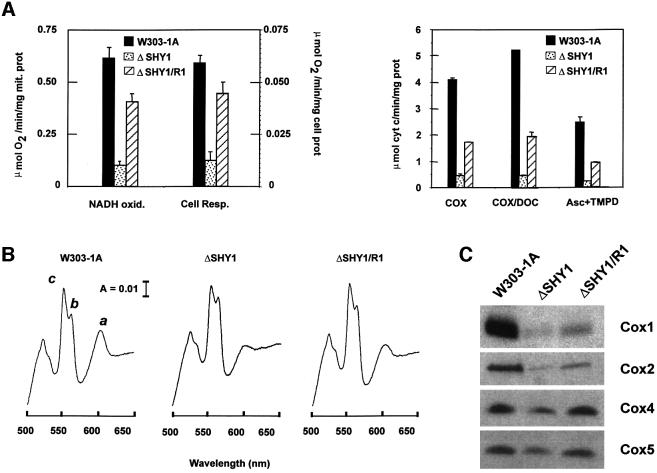
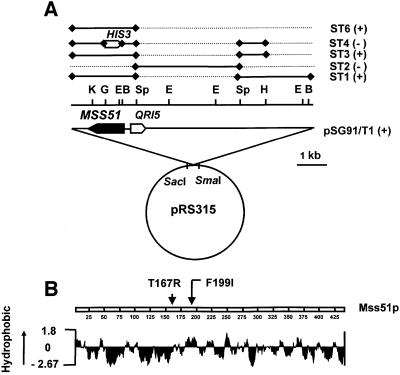
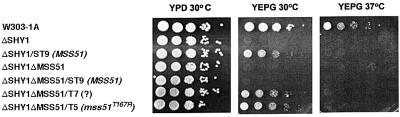
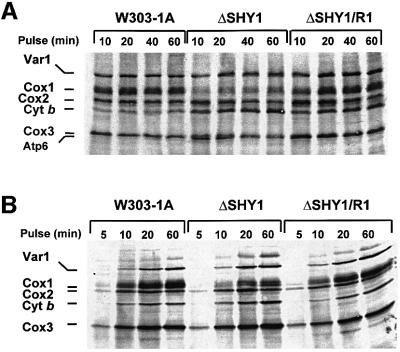
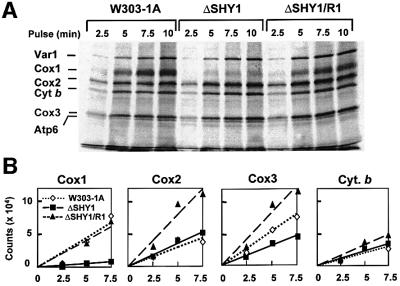
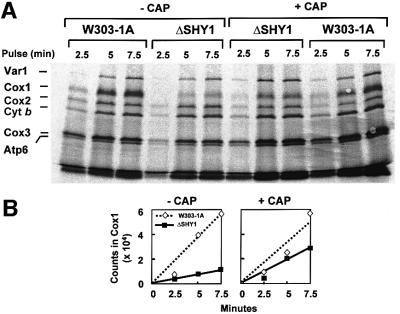
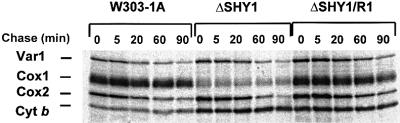
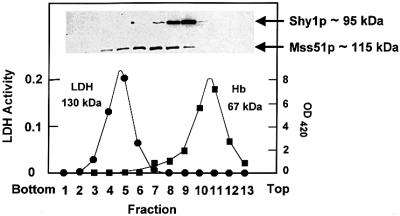
SHY1 codes for a mitochondrial protein required for full expression of cytochrome oxidase (COX) in Saccharomyces cerevisiae. Mutations in the homologous human gene (SURF1) have been reported to cause Leigh's syndrome, a neurological disease associated with COX deficiency. The function of Shy1p/Surf1p is poorly understood. Here we have characterized revertants of shy1 null mutants carrying extragenic nuclear suppressor mutations. The steady-state levels of COX in the revertants is increased by a factor of 4-5, accounting for their ability to respire and grow on non-fermentable carbon sources at nearly wild-type rates. The suppressor mutations are in MSS51, a gene previously implicated in processing and translation of the COX1 transcript for subunit 1 (Cox1) of COX. The function of Shy1p and the mechanism of suppression of shy1 mutants were examined by comparing the rates of synthesis and turnover of the mitochondrial translation products in wild-type, mutant and revertant cells. We propose that Shy1p promotes the formation of an assembly intermediate in which Cox1 is one of the partners.
Figures
References
-
- Beyer R.E. (1964) A protein factor required for phosphorylation coupled to electron flow between reduced coenzyme Q and cytochrome c in the electron transfer chain. Biochem. Biophys. Res. Commun., 16, 460–465. - PubMed
-
- Choquet Y., Wostrikoff,K., Rimbault,B., Zito,F., Girard-Bascou,J., Drapier,D. and Wollman,F.A. (2001) Assembly-controlled regulation of chloroplast gene translation. Biochem Soc. Trans., 29, 421–426. - PubMed
-
- Coenen M.J., van den Heuvel,L.P., Nijtmans,L.G., Morava,E., Marquardt,I., Girschick,H.J., Trijbels,F.J., Grivell,L.A. and Smeitink, J.A. (1999) SURFEIT-1 gene analysis and two-dimensional blue native gel electrophoresis in cytochrome c oxidase deficiency. Biochem. Biophys. Res. Commun., 265, 339–344. - PubMed
-
- Decoster E., Simon,M., Hatat,D. and Faye,G. (1990) The MSS51 gene product is required for the translation of the COX1 mRNA in yeast mitochondria. Mol. Gen. Genet., 224, 111–118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
