

Regulation of Raf-1 activation and signalling by dephosphorylation
- PMID: 11782426
- PMCID: PMC125807
- DOI: 10.1093/emboj/21.1.64
Regulation of Raf-1 activation and signalling by dephosphorylation
Abstract
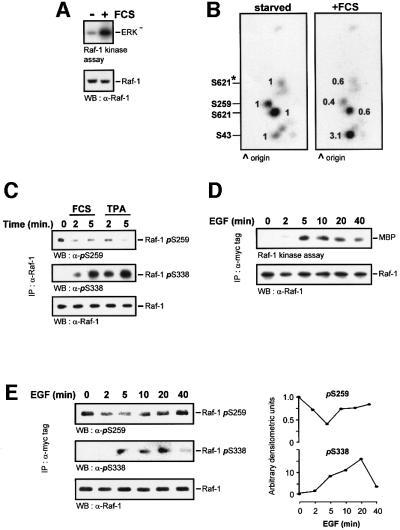
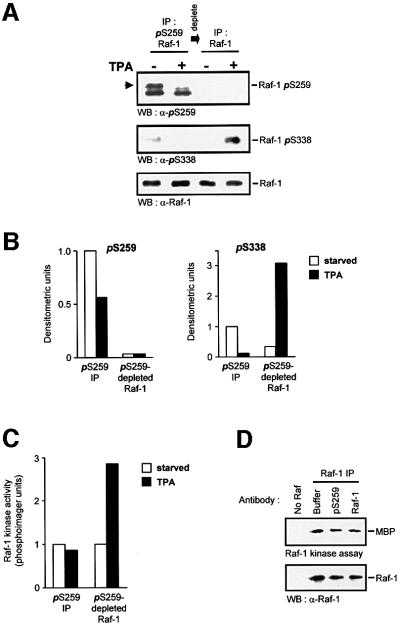
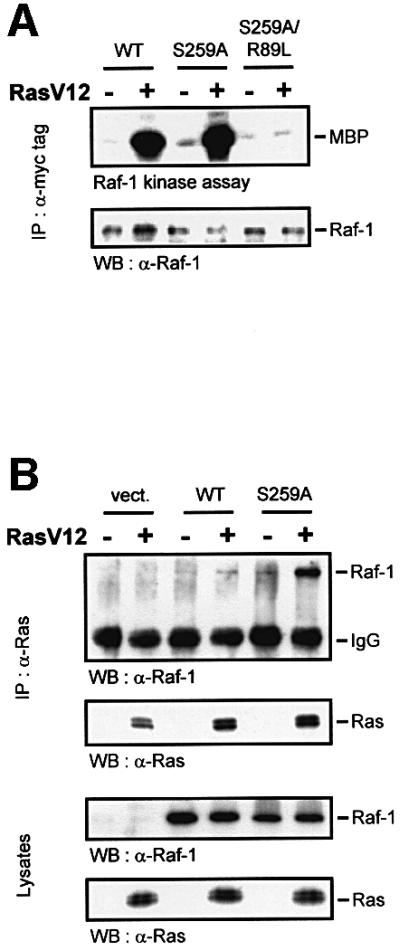
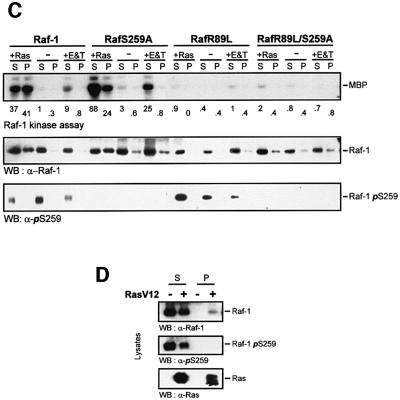
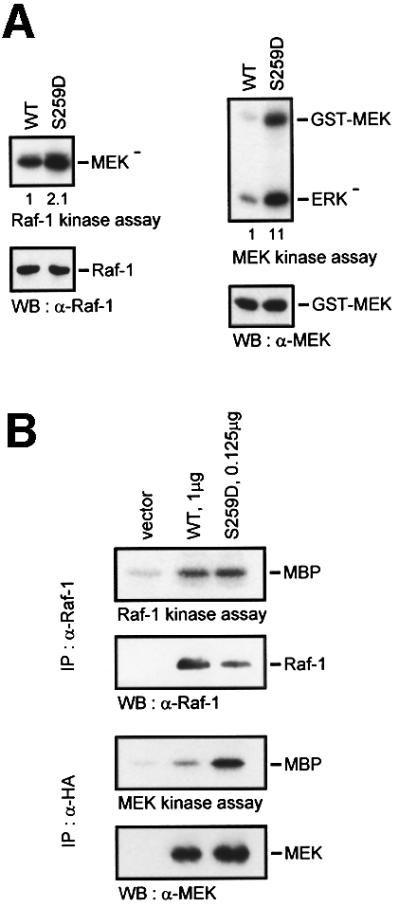
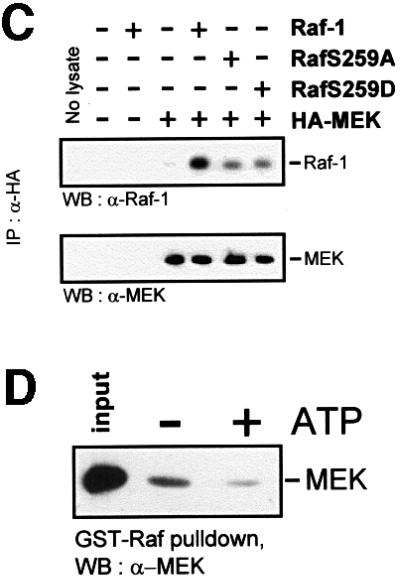
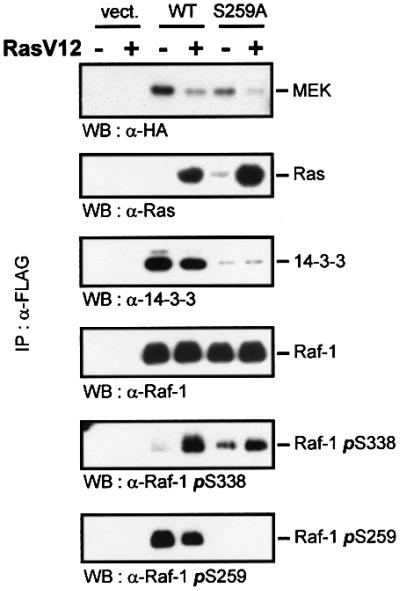
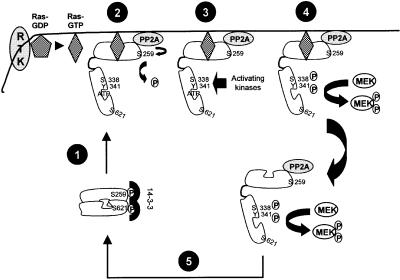
The Raf-1 kinase is regulated by phosphorylation, and Ser259 has been identified as an inhibitory phosphorylation site. Here we show that the dephosphorylation of Ser259 is an essential part of the Raf-1 activation process, and further reveal the molecular role of Ser259. The fraction of Raf-1 that is phosphorylated on Ser259 is refractory to mitogenic stimulation. Mutating Ser259 elevates kinase activity because of enhanced binding to Ras and constitutive membrane recruitment. This facilitates the phosphorylation of an activating site, Ser338. The mutation of Ser259 also increases the functional coupling to MEK, augmenting the efficiency of MEK activation. Our results suggest that Ser259 regulates the coupling of Raf-1 to upstream activators as well as to its downstream substrate MEK, thus determining the pool of Raf-1 that is competent for signalling. They also suggest a new model for Raf-1 activation where the release of repression through Ser259 dephosphorylation is the pivotal step.
Figures
References
-
- Abraham D. et al. (2000) Raf-1-associated protein phosphatase 2A as a positive regulator of kinase activation. J. Biol. Chem., 275, 22300–22304. - PubMed
-
- Alessandrini A., Greulich,H., Huang,W.D. and Erikson,R.L. (1996) Mek1 phosphorylation site mutants activate Raf-1 in NIH 3T3 cells. J. Biol. Chem., 271, 31612–31618. - PubMed
-
- Alessi D.R., Cuenda,A., Cohen,P., Dudley,D.T. and Saltiel,A.R. (1995) PD 098059 is a specific inhibitor of the activation of mitogen-activated protein kinase kinase in vitro and in vivo. J. Biol. Chem., 270, 27489–27494. - PubMed
-
- Avruch J., Khokhlatchev,A., Kyriakis,J.M., Luo,Z., Tzivion,G., Vavvas, D. and Zhang,X.F. (2001) Ras activation of the Raf kinase: tyrosine kinase recruitment of the MAP kinase cascade. Recent Prog. Horm. Res., 56, 127–155. - PubMed
-
- Barnard D., Diaz,B., Clawson,D. and Marshall,M. (1998) Oncogenes, growth factors and phorbol esters regulate Raf-1 through common mechanisms. Oncogene, 17, 1539–1547. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
