

A novel RNA polymerase I transcription initiation factor, TIF-IE, commits rRNA genes by interaction with TIF-IB, not by DNA binding
- PMID: 11784852
- PMCID: PMC133551
- DOI: 10.1128/MCB.22.3.750-761.2002
A novel RNA polymerase I transcription initiation factor, TIF-IE, commits rRNA genes by interaction with TIF-IB, not by DNA binding
Abstract
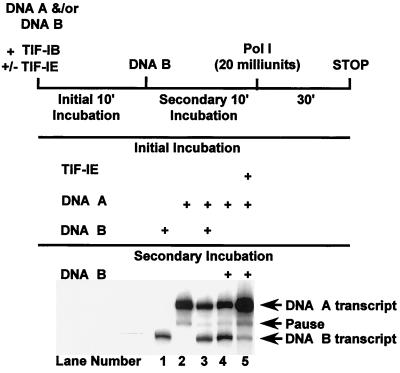
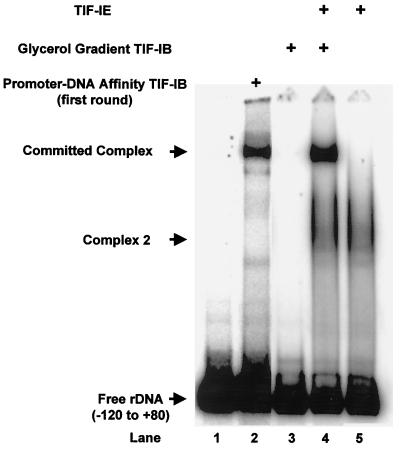
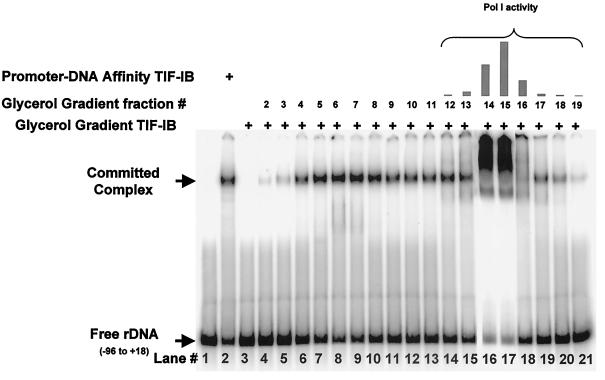
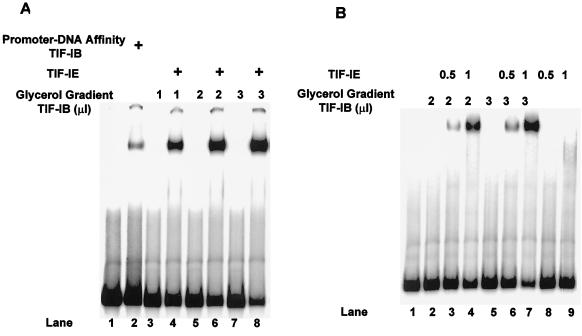
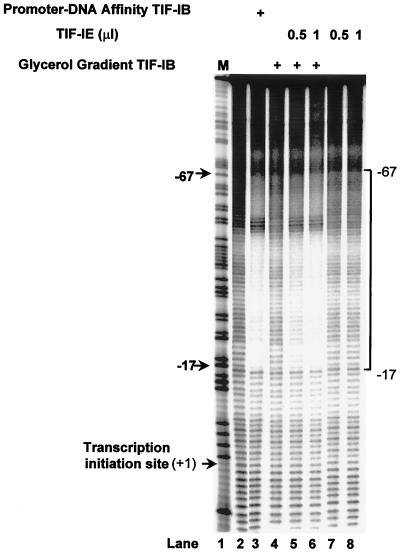
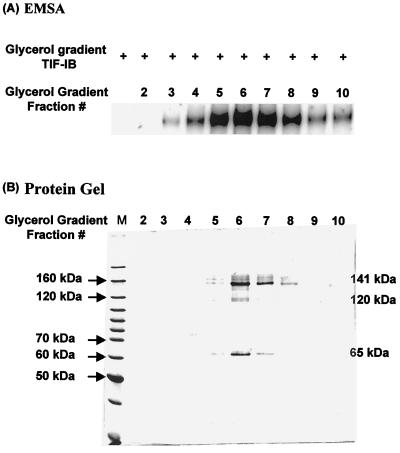
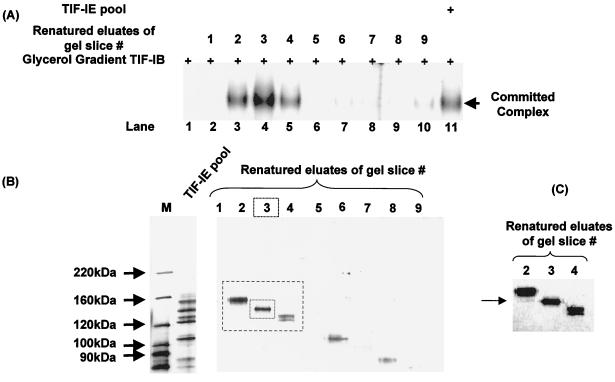
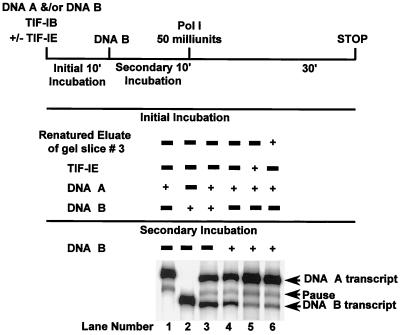
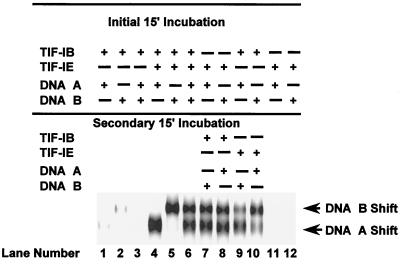
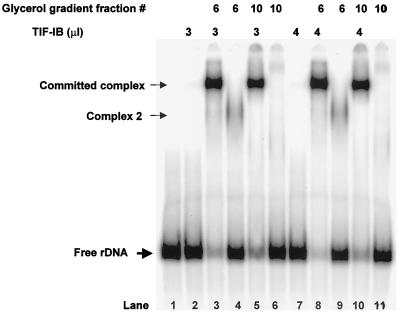
In the small, free-living amoeba Acanthamoeba castellanii, rRNA transcription requires, in addition to RNA polymerase I, a single DNA-binding factor, transcription initiation factor IB (TIF-IB). TIF-IB is a multimeric protein that contains TATA-binding protein (TBP) and four TBP-associated factors that are specific for polymerase I transcription. TIF-IB is required for accurate and promoter-specific initiation of rRNA transcription, recruiting and positioning the polymerase on the start site by protein-protein interaction. In A. castellanii, partially purified TIF-IB can form a persistent complex with the ribosomal DNA (rDNA) promoter while homogeneous TIF-IB cannot. An additional factor, TIF-IE, is required along with homogeneous TIF-IB for the formation of a stable complex on the rDNA core promoter. We show that TIF-IE by itself, however, does not bind to the rDNA promoter and thus differs in its mechanism from the upstream binding factor and upstream activating factor, which carry out similar complex-stabilizing functions in vertebrates and yeast, respectively. In addition to its presence in impure TIF-IB, TIF-IE is found in highly purified fractions of polymerase I, with which it associates. Renaturation of polypeptides excised from sodium dodecyl sulfate-polyacrylamide gels showed that a 141-kDa polypeptide possesses all the known activities of TIF-IE.
Figures

References
-
- Bachvarov, D., M. Normandeau, and T. Moss. 1991. Heterogeneity in the Xenopus ribosomal transcription factor xUBF has a molecular basis distinct from that in mammals. FEBS Lett. 288:55–59. - PubMed
-
- Bateman, E., L. Hoffman, C. Iida, W. Kubaska, P. Kownin, P. Risi, M. Zwick, and M. R. Paule. 1989. Eukaryotic RNA polymerase I binds to promoters by protein-protein interactions and melts DNA in a separate step following binding, p.259–269. In T. Cech and J. Gralla (ed.), Molecular biology of RNA- and DNA-protein interactions in transcription. Alan R. Liss, Inc., New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources