

Retinoblastoma protein transcriptional repression through histone deacetylation of a single nucleosome
- PMID: 11784861
- PMCID: PMC133558
- DOI: 10.1128/MCB.22.3.856-865.2002
Retinoblastoma protein transcriptional repression through histone deacetylation of a single nucleosome
Abstract
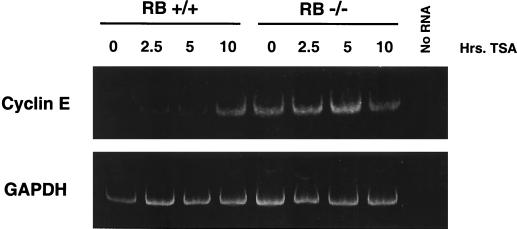
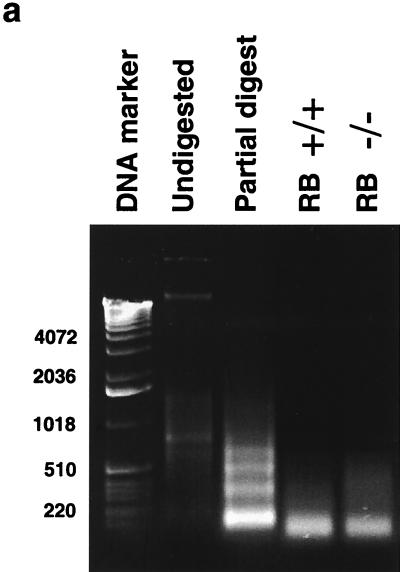
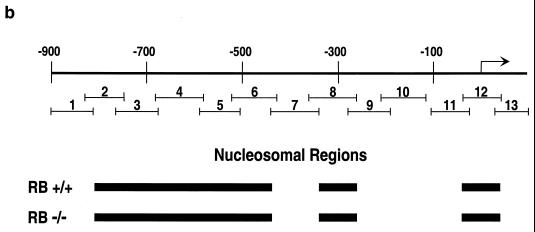
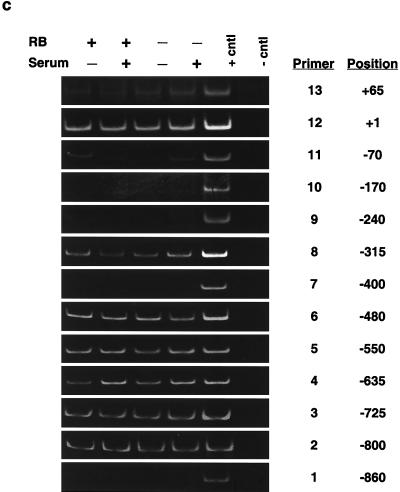
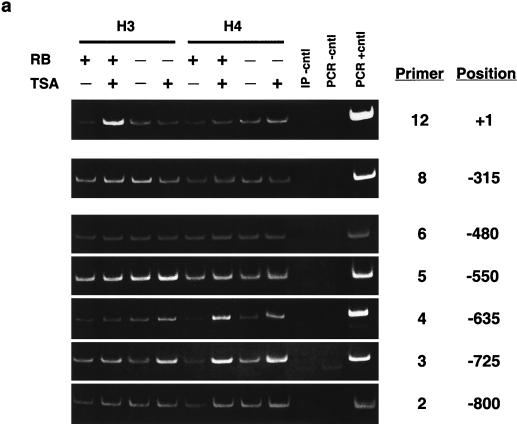
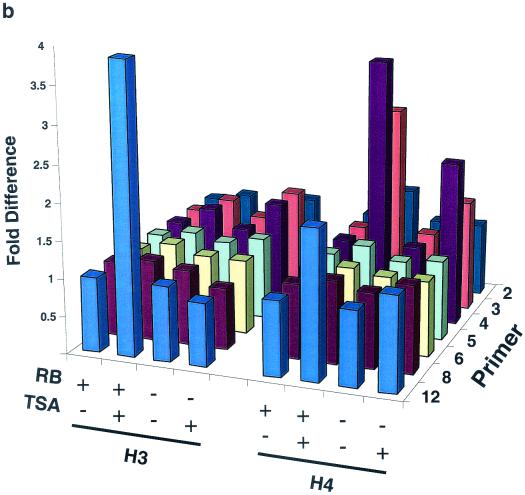
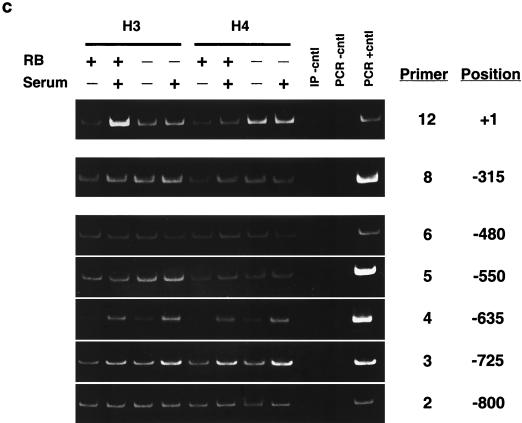
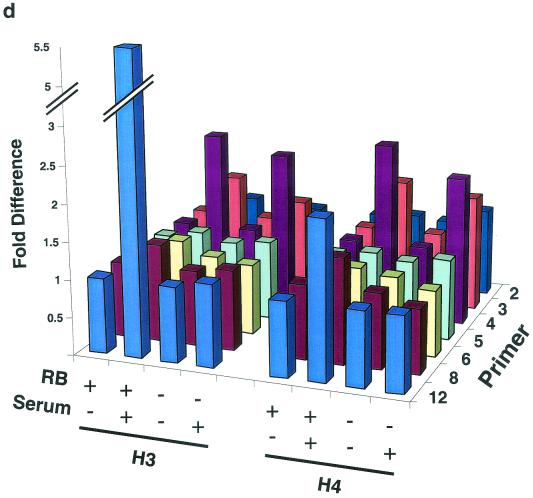
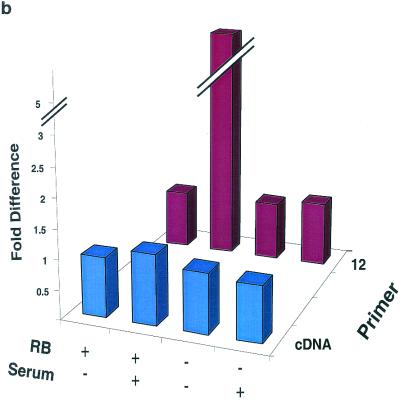
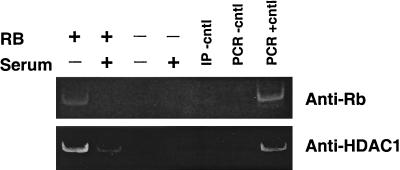
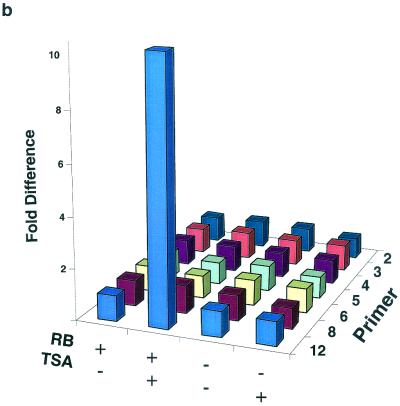
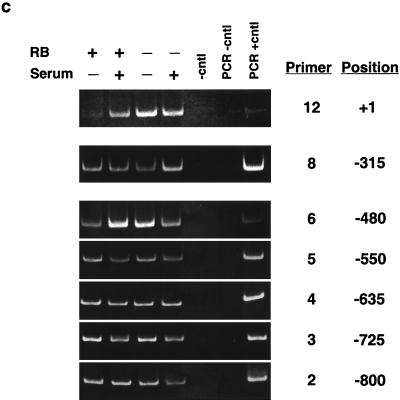
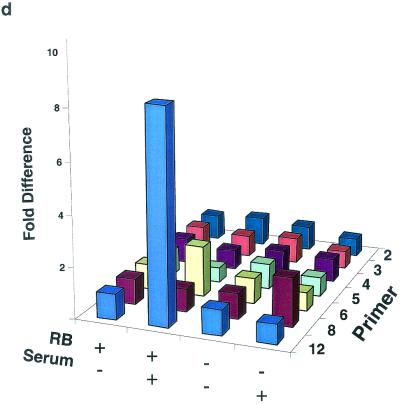
The retinoblastoma protein, pRb, controls transcription through recruitment of histone deacetylase to particular E2F-responsive genes. We determined the acetylation level of individual nucleosomes present in the cyclin E promoter of RB(+/+) and RB(-/-) mouse embryo fibroblasts. We also determined the effects of pRb on nucleosomal conformation by examining the thiol reactivity of histone H3 of individual nucleosomes. We found that pRb represses the cyclin E promoter through histone deacetylation of a single nucleosome, to which it and histone deacetylase 1 bind. In addition, the conformation of this nucleosome is modulated by pRb-directed histone deacetylase activity. Thus, the repressive role of pRb in cyclin E transcription and therefore cell cycle progression can be mapped to its control of the acetylation status and conformation of a single nucleosome.
Figures


Similar articles
-
Roles of pRB in the Regulation of Nucleosome and Chromatin Structures.Biomed Res Int. 2016;2016:5959721. doi: 10.1155/2016/5959721. Epub 2016 Dec 22. Biomed Res Int. 2016. PMID: 28101510 Free PMC article. Review.
-
E2F mediates cell cycle-dependent transcriptional repression in vivo by recruitment of an HDAC1/mSin3B corepressor complex.Genes Dev. 2002 Apr 15;16(8):933-47. doi: 10.1101/gad.969202. Genes Dev. 2002. PMID: 11959842 Free PMC article.
-
Histone deacetylation of RB-responsive promoters: requisite for specific gene repression but dispensable for cell cycle inhibition.Mol Cell Biol. 2003 Nov;23(21):7719-31. doi: 10.1128/MCB.23.21.7719-7731.2003. Mol Cell Biol. 2003. PMID: 14560017 Free PMC article.
-
Epigenetic control of mammalian LINE-1 retrotransposon by retinoblastoma proteins.Mutat Res. 2009 Jun 1;665(1-2):20-8. doi: 10.1016/j.mrfmmm.2009.02.011. Epub 2009 Mar 9. Mutat Res. 2009. PMID: 19427507 Free PMC article.
-
Histone acetylation and the cell-cycle in cancer.Front Biosci. 2001 Apr 1;6:D610-29. doi: 10.2741/1wang1. Front Biosci. 2001. PMID: 11282573 Review.
Cited by
-
Retinoblastoma Tumor Suppressor Protein Roles in Epigenetic Regulation.Cancers (Basel). 2020 Sep 29;12(10):2807. doi: 10.3390/cancers12102807. Cancers (Basel). 2020. PMID: 33003565 Free PMC article. Review.
-
Regulation of transcription and chromatin structure by pRB: here, there and everywhere.Cell Cycle. 2012 Sep 1;11(17):3189-98. doi: 10.4161/cc.21263. Epub 2012 Aug 16. Cell Cycle. 2012. PMID: 22895179 Free PMC article. Review.
-
The p53 regulatory gene MDM2 is a direct transcriptional target of MYCN in neuroblastoma.Proc Natl Acad Sci U S A. 2005 Jan 18;102(3):731-6. doi: 10.1073/pnas.0405495102. Epub 2005 Jan 11. Proc Natl Acad Sci U S A. 2005. PMID: 15644444 Free PMC article.
-
Roles of pRB in the Regulation of Nucleosome and Chromatin Structures.Biomed Res Int. 2016;2016:5959721. doi: 10.1155/2016/5959721. Epub 2016 Dec 22. Biomed Res Int. 2016. PMID: 28101510 Free PMC article. Review.
-
p65 Negatively regulates transcription of the cyclin E gene.J Biol Chem. 2010 Jun 4;285(23):17453-64. doi: 10.1074/jbc.M109.058974. Epub 2010 Apr 11. J Biol Chem. 2010. PMID: 20385564 Free PMC article.
References
-
- Agalioti, T., S. Lomvardas, B. Parekh, J. Yie, T. Maniatis, and D. Thanos. 2000. Ordered recruitment of chromatin modifying and general transcription factors to the IFN-beta promoter. Cell 103:667–678. - PubMed
-
- Allegra, P., R. Sterner, D. F. Clayton, and V. G. Allfrey. 1987. Affinity chromatographic purification of nucleosomes containing transcriptionally active DNA sequences. J. Mol. Biol. 196:379–388. - PubMed
-
- Boffa, L. C., J. Walker, T. A. Chen, R. Sterner, M. R. Mariani, and V. G. Allfrey. 1990. Factors affecting nucleosome structure in transcriptionally active chromatin. Histone acetylation, nascent RNA and inhibitors of RNA synthesis. Eur. J. Biochem. 194:811–823. - PubMed
-
- Boggs, B. A., B. Connors, R. E. Sobel, A. C. Chinault, and C. D. Allis. 1996. Reduced levels of histone H3 acetylation on the inactive X chromosome in human females. Chromosoma 105:303–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources