

Molecular balance between the regulatory and catalytic subunits of phosphoinositide 3-kinase regulates cell signaling and survival
- PMID: 11784871
- PMCID: PMC133541
- DOI: 10.1128/MCB.22.3.965-977.2002
Molecular balance between the regulatory and catalytic subunits of phosphoinositide 3-kinase regulates cell signaling and survival
Abstract
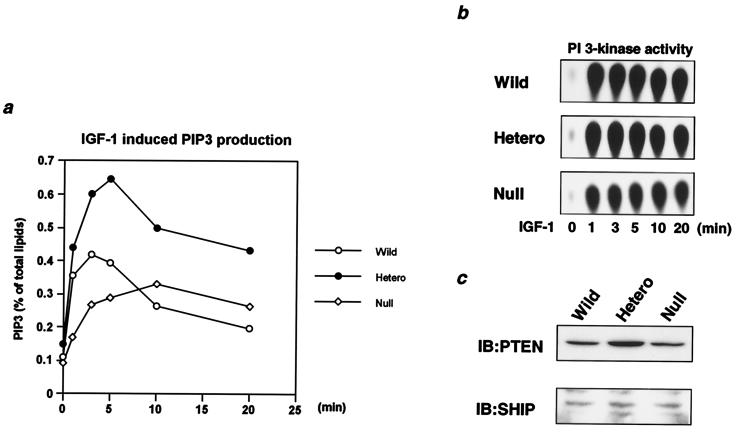
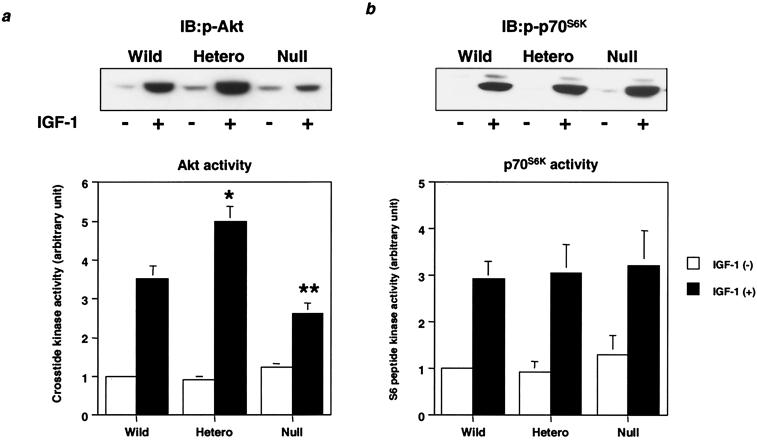
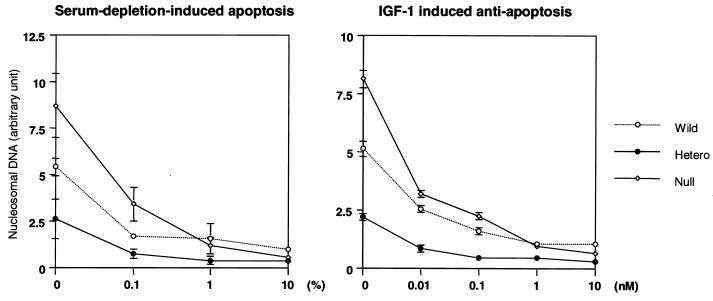
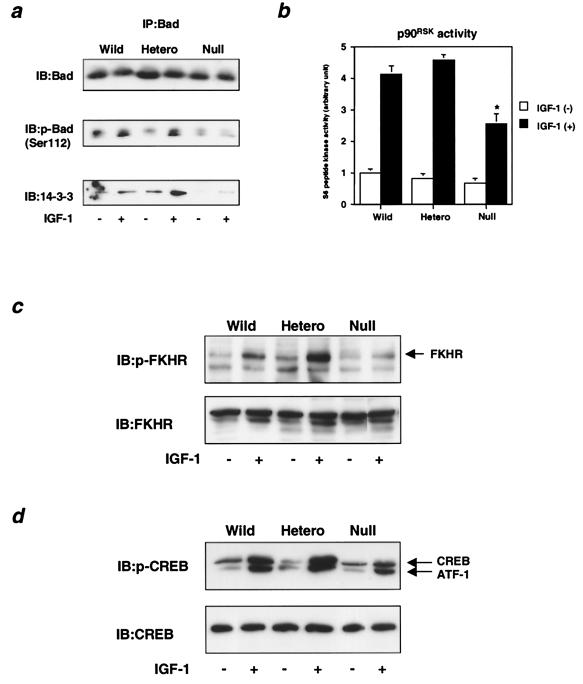
Class Ia phosphoinositide (PI) 3-kinase is a central component in growth factor signaling and is comprised of a p110 catalytic subunit and a regulatory subunit, the most common family of which is derived from the p85alpha gene (Pik3r1). Optimal signaling through the PI 3-kinase pathway depends on a critical molecular balance between the regulatory and catalytic subunits. In wild-type cells, the p85 subunit is more abundant than p110, leading to competition between the p85 monomer and the p85-p110 dimer and ineffective signaling. Heterozygous disruption of Pik3r1 results in increased Akt activity and decreased apoptosis by insulin-like growth factor 1 (IGF-1) through up-regulated phosphatidylinositol (3,4,5)-triphosphate production. Complete depletion of p85alpha, on the other hand, results in significantly increased apoptosis due to reduced PI 3-kinase-dependent signaling. Thus, a reduction in p85alpha represents a novel therapeutic target for enhancing IGF-1/insulin signaling, prolongation of cell survival, and protection against apoptosis.
Figures
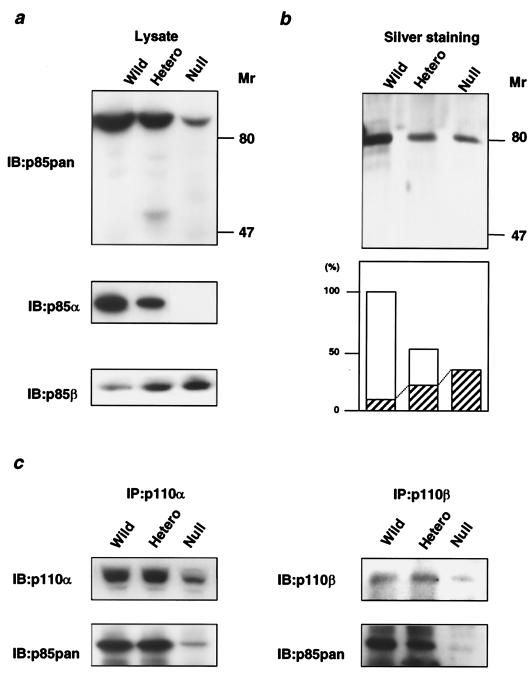
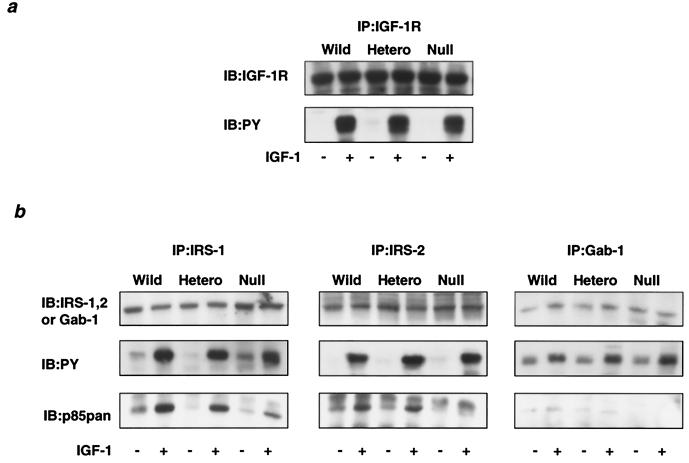
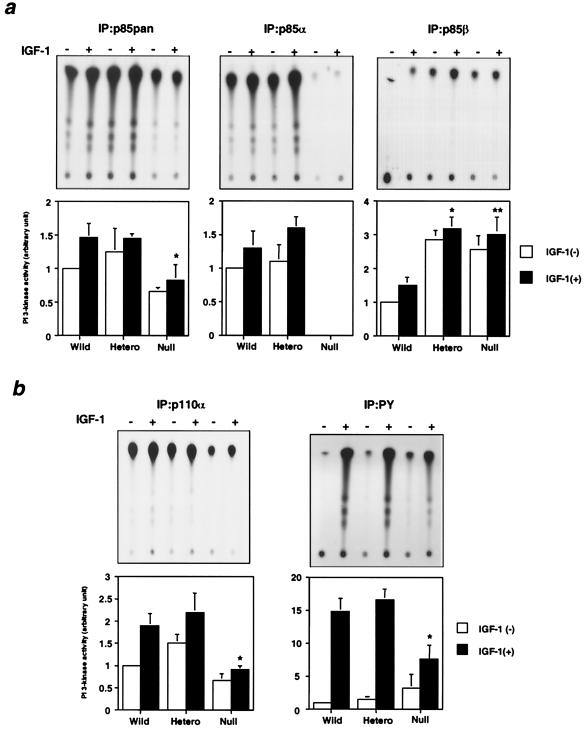
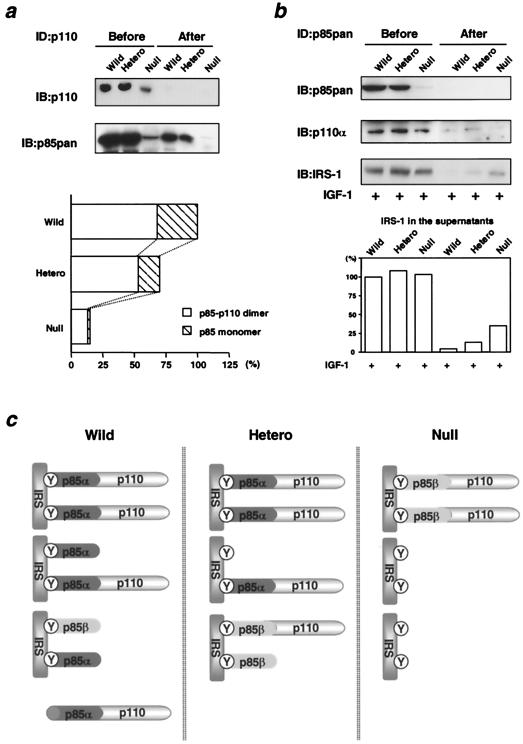
References
-
- Baltensperger, K., L. M. Kozma, S. R. Jaspers, and M. P. Czech. 1994. Regulation by insulin of phosphatidylinositol 3′-kinase bound to alpha- and beta-isoforms of p85 regulatory subunit. J. Biol. Chem. 269:28937–28946. - PubMed
-
- Bonni, A., A. Brunet, A. E. West, S. R. Datta, M. A. Takasu, and M. E. Greenberg. 1999. Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 286:1358–1362. - PubMed
-
- Brunet, A., A. Bonni, M. J. Zigmond, M. Z. Lin, P. Juo, L. S. Hu, M. J. Anderson, K. C. Arden, J. Blenis, and M. E. Greenberg. 1999. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 96:857–868. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous