

Bone morphogenetic protein-2 (BMP-2) transactivates Dlx3 through Smad1 and Smad4: alternative mode for Dlx3 induction in mouse keratinocytes
- PMID: 11788714
- PMCID: PMC99823
- DOI: 10.1093/nar/30.2.515
Bone morphogenetic protein-2 (BMP-2) transactivates Dlx3 through Smad1 and Smad4: alternative mode for Dlx3 induction in mouse keratinocytes
Abstract
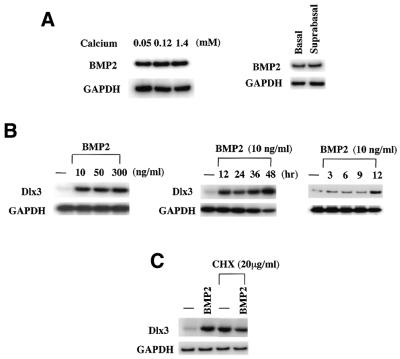
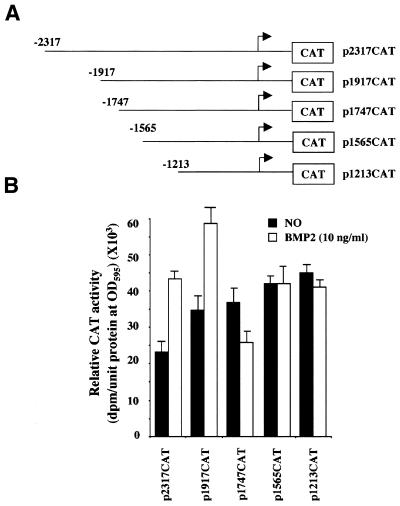
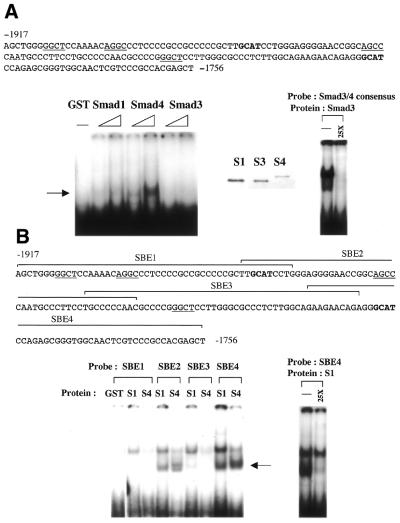
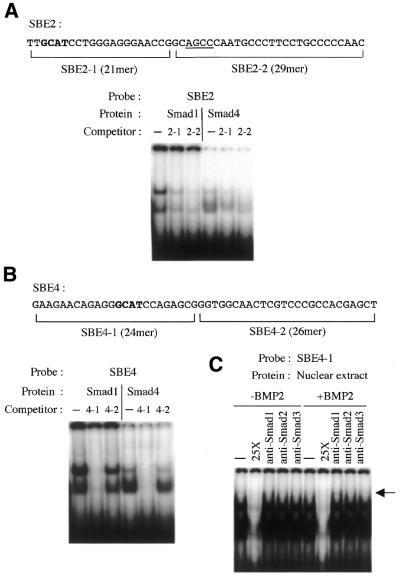
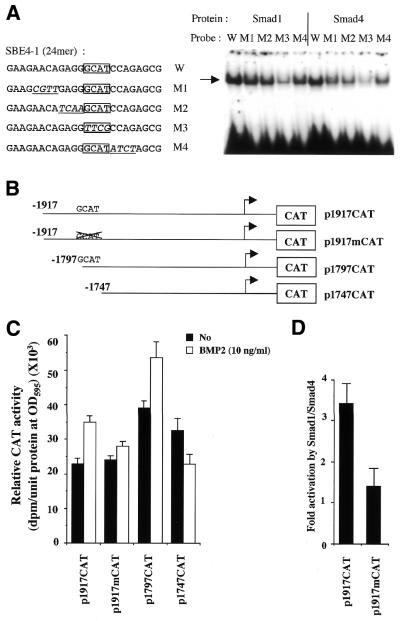
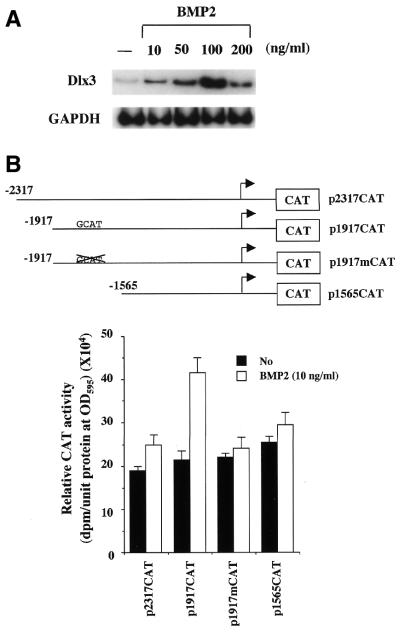
Expression of the Dlx3 homeodomain gene is induced in terminally differentiated epidermal cells. Dlx3 regulates gene expression in skin and plays important roles in patterning of the embryonic ectoderm through differential sensitivity to bone morphogenetic protein (BMP) signaling. We analyzed the expression of BMP family members in murine keratinocytes; BMP-2 is expressed in proliferative basal and differentiated suprabasal keratinocytes. BMP-2 induced transcription of Dlx3 within 12 h of treatment of keratinocytes cultured in vitro. We proceeded to delineate the BMP-2-responsive region to an area between -1917 and -1747 in the Dlx3 promoter. Gel shift assays with recombinant Smad1 and Smad4 demonstrated that this DNA fragment (-1917 to -1747) was competent in the formation of protein-DNA complexes. By deletion and mutational analyses we localized a Smad1/Smad4-binding site containing a GCAT motif, which showed similarity to other TGF-beta family responsive elements. Supershift assays with keratinocyte nuclear extracts and antibodies against members of the Smad family showed that this motif was able to form a complex with Smad1. Mutation of the Smad1/Smad4-binding site inhibited transcriptional activation of the Dlx3 gene by BMP-2. In the hair follicle, where Dlx3 is expressed in the hair matrix cells, BMP-2 also activates Dlx3 transcription. These results provide a possible mechanism of action for the BMP signaling pathway on the regulation of Dlx3.
Figures
Similar articles
-
Smad1 and Smad4 are components of the bone morphogenetic protein-4 (BMP-4)-induced transcription complex of the Xvent-2B promoter.J Biol Chem. 2000 Jul 21;275(29):21827-35. doi: 10.1074/jbc.M000978200. J Biol Chem. 2000. PMID: 10791953
-
Characterization of a bone morphogenetic protein-responsive Smad-binding element.Mol Biol Cell. 2000 Feb;11(2):555-65. doi: 10.1091/mbc.11.2.555. Mol Biol Cell. 2000. PMID: 10679014 Free PMC article.
-
Direct binding of Smad1 and Smad4 to two distinct motifs mediates bone morphogenetic protein-specific transcriptional activation of Id1 gene.J Biol Chem. 2002 Feb 1;277(5):3176-85. doi: 10.1074/jbc.M106826200. Epub 2001 Nov 7. J Biol Chem. 2002. PMID: 11700304
-
Bone morphogenetic proteins.Growth Factors. 2004 Dec;22(4):233-41. doi: 10.1080/08977190412331279890. Growth Factors. 2004. PMID: 15621726 Review.
-
Bone Morphogenetic Proteins.Cold Spring Harb Perspect Biol. 2016 Jun 1;8(6):a021899. doi: 10.1101/cshperspect.a021899. Cold Spring Harb Perspect Biol. 2016. PMID: 27252362 Free PMC article. Review.
Cited by
-
Msx and dlx homeogene expression in epithelial odontogenic tumors.J Histochem Cytochem. 2009 Jan;57(1):69-78. doi: 10.1369/jhc.2008.951707. Epub 2008 Oct 14. J Histochem Cytochem. 2009. PMID: 18854600 Free PMC article.
-
Homeodomain protein DLX4 counteracts key transcriptional control mechanisms of the TGF-β cytostatic program and blocks the antiproliferative effect of TGF-β.Oncogene. 2011 Jun 16;30(24):2718-29. doi: 10.1038/onc.2011.4. Epub 2011 Feb 7. Oncogene. 2011. PMID: 21297662 Free PMC article.
-
Distal-less 3 haploinsufficiency results in elevated placental oxidative stress and altered fetal growth kinetics in the mouse.Placenta. 2012 Oct;33(10):830-8. doi: 10.1016/j.placenta.2012.06.018. Epub 2012 Jul 19. Placenta. 2012. PMID: 22819041 Free PMC article.
-
Preliminary study on microR-148a and microR-10a in dermal papilla cells of Hu sheep.BMC Genet. 2019 Aug 27;20(1):70. doi: 10.1186/s12863-019-0770-8. BMC Genet. 2019. PMID: 31455210 Free PMC article.
-
An Oct-1 binding site mediates activation of the gata2 promoter by BMP signaling.Nucleic Acids Res. 2005 Aug 1;33(13):4357-67. doi: 10.1093/nar/gki746. Print 2005. Nucleic Acids Res. 2005. PMID: 16061939 Free PMC article.
References
-
- Fuchs E. and Byrne,C. (1994) The epidermis: rising to the surface. Curr. Opin. Genet. Dev., 4, 725–736. - PubMed
-
- Price J.A., Bowden,D.W., Wright,J.T., Pettenati,M.J. and Hart,T.C. (1998) Identification of a mutation in DLX3 associated with tricho-dento-osseous (TDO) syndrome. Hum. Mol. Genet., 7, 563–569. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
