

Phytochrome E controls light-induced germination of Arabidopsis
- PMID: 11788765
- PMCID: PMC148973
Phytochrome E controls light-induced germination of Arabidopsis
Abstract
Germination of Arabidopsis seeds is light dependent and under phytochrome control. Previously, phytochromes A and B and at least one additional, unspecified phytochrome were shown to be involved in this process. Here, we used a set of photoreceptor mutants to test whether phytochrome D and/or phytochrome E can control germination of Arabidopsis. The results show that only phytochromes B and E, but not phytochrome D, participate directly in red/far-red light (FR)-reversible germination. Unlike phytochromes B and D, phytochrome E did not inhibit phytochrome A-mediated germination. Surprisingly, phytochrome E was required for germination of Arabidopsis seeds in continuous FR. However, inhibition of hypocotyl elongation by FR, induction of cotyledon unfolding, and induction of agravitropic growth were not affected by loss of phytochrome E. Therefore, phytochrome E is not required per se for phytochrome A-mediated very low fluence responses and the high irradiance response. Immunoblotting revealed that the need of phytochrome E for germination in FR was not caused by altered phytochrome A levels. These results uncover a novel role of phytochrome E in plant development and demonstrate the considerable functional diversification of the closely related phytochromes B, D, and E.
Figures
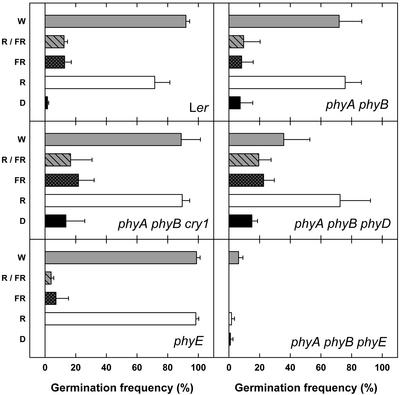
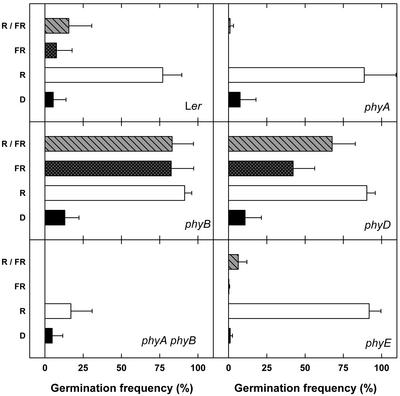
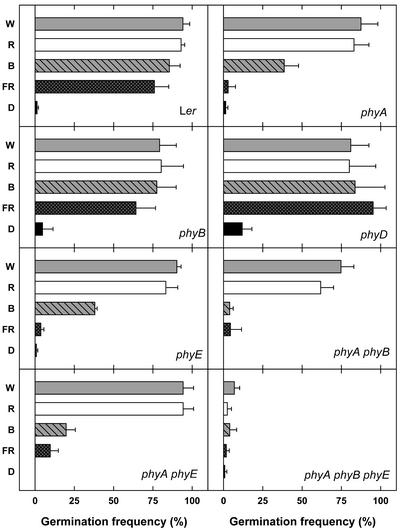
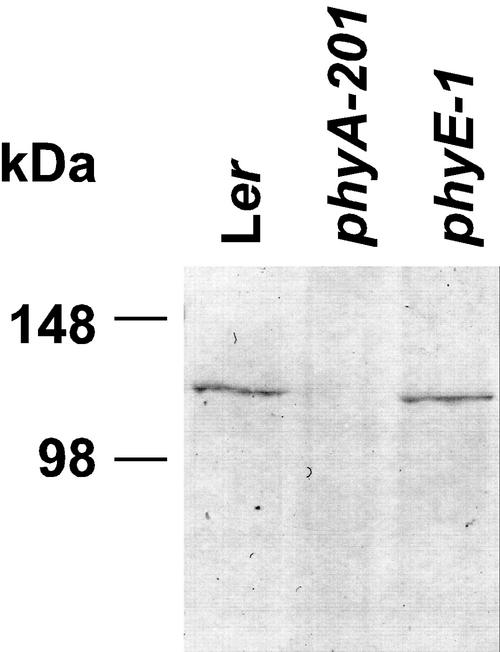
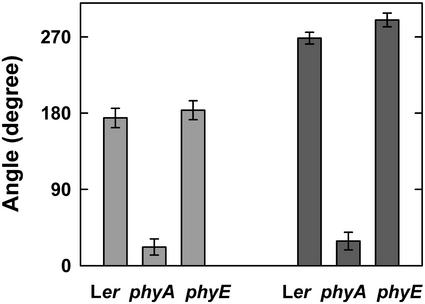
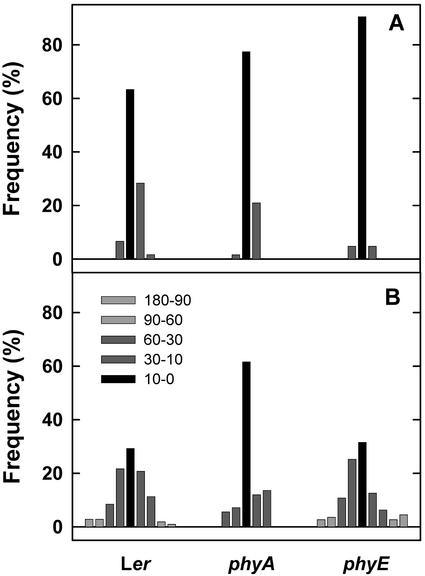
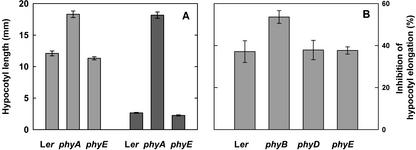
References
-
- Ahmad M, Cashmore AR. HY4 gene of Arabidopsis thaliana encodes a protein with characteristics of a blue-light photoreceptor. Nature. 1993;366:162–166. - PubMed
-
- Casal JJ. Phytochromes, cryptochromes, phototropin: photoreceptor interactions in plants. Photochem Photobiol. 2000;71:1–11. - PubMed
-
- Casal JJ, Sánchez RA. Phytochromes and seed germination. Seed Sci Res. 1998;8:317–329.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources