

Identification of the periplasmic cobalamin-binding protein BtuF of Escherichia coli
- PMID: 11790740
- PMCID: PMC139523
- DOI: 10.1128/JB.184.3.706-717.2002
Identification of the periplasmic cobalamin-binding protein BtuF of Escherichia coli
Abstract
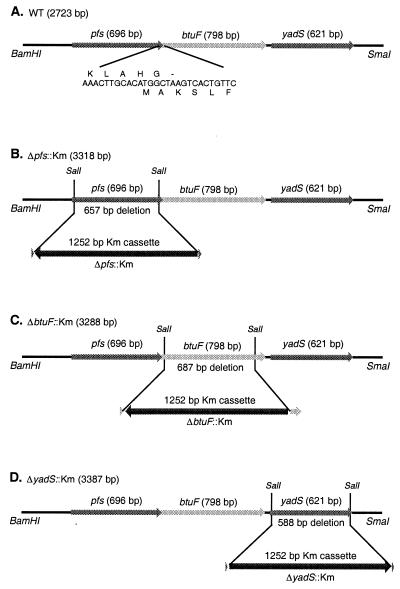
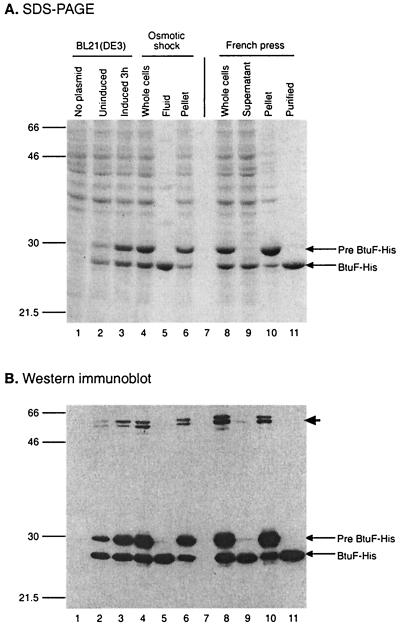
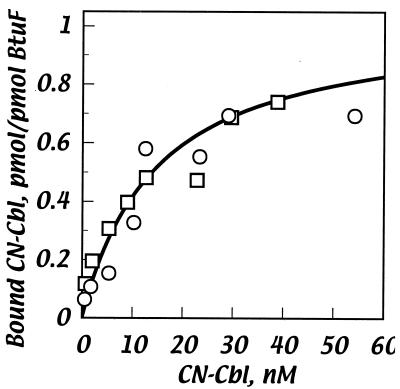
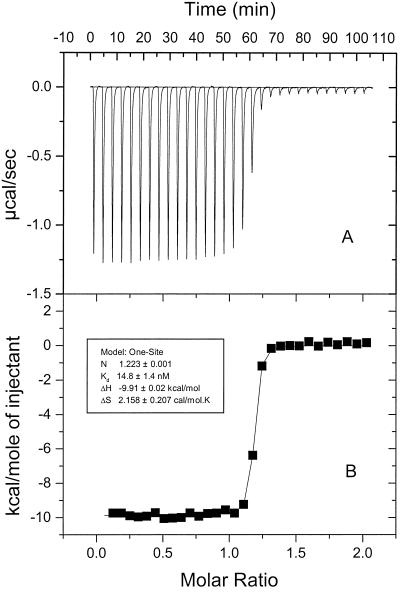
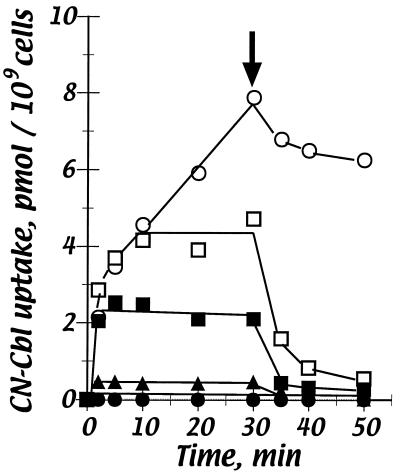
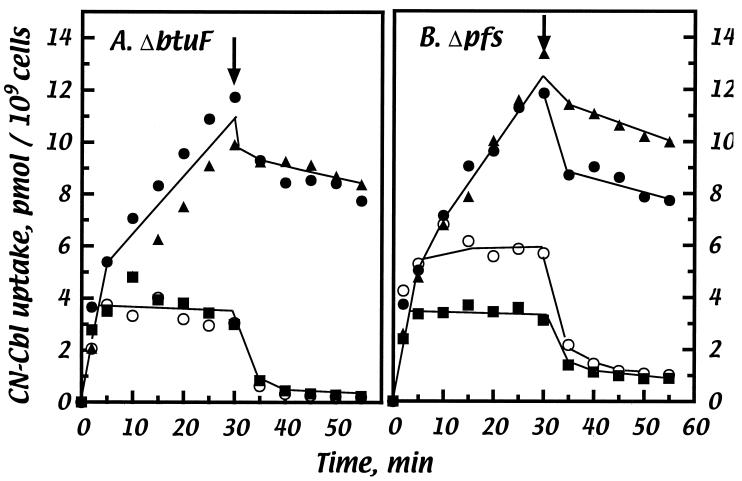
Cells of Escherichia coli take up vitamin B(12) (cyano-cobalamin [CN-Cbl]) and iron chelates by use of sequential active transport processes. Transport of CN-Cbl across the outer membrane and its accumulation in the periplasm is mediated by the TonB-dependent transporter BtuB. Transport across the cytoplasmic membrane (CM) requires the BtuC and BtuD proteins, which are most related in sequence to the transmembrane and ATP-binding cassette proteins of periplasmic permeases for iron-siderophore transport. Unlike the genetic organization of most periplasmic permeases, a candidate gene for a periplasmic Cbl-binding protein is not linked to the btuCED operon. The open reading frame termed yadT in the E. coli genomic sequence is related in sequence to the periplasmic binding proteins for iron-siderophore complexes and was previously implicated in CN-Cbl uptake in Salmonella. The E. coli yadT product, renamed BtuF, is shown here to participate in CN-Cbl uptake. BtuF protein, expressed with a C-terminal His(6) tag, was shown to be translocated to the periplasm concomitant with removal of a signal sequence. CN-Cbl-binding assays using radiolabeled substrate or isothermal titration calorimetry showed that purified BtuF binds CN-Cbl with a binding constant of around 15 nM. A null mutation in btuF, but not in the flanking genes pfs and yadS, strongly decreased CN-Cbl utilization and transport into the cytoplasm. The growth response to CN-Cbl of the btuF mutant was much stronger than the slight impairment previously described for btuC, btuD, or btuF mutants. Hence, null mutations in btuC and btuD were constructed and revealed that the btuC mutant had a strong impairment similar to that of the btuF mutant, whereas the btuD defect was less pronounced. All mutants with defective transport across the CM gave rise to frequent suppressor variants which were able to respond at lower levels of CN-Cbl but were still defective in transport across the CM. These results finally establish the identity of the periplasmic binding protein for Cbl uptake, which is one of few cases where the components of a periplasmic permease are genetically separated.
Figures
References
-
- Berlyn, M. K. B., K. B. Low, and K. E. Rudd. 1996. Linkage map of Escherichia coli K-12, edition 9, p.1715–1902. In F. C. Neidhardt, R. Curtiss III, J. L. Ingraham, E. C. C. Lin, K. B. Low, B. Magasanik, W. E. Reznikoff, M. Riley, M. Schaechter, and H. E. Umbarger (ed.), Escherichia coli and Salmonella: cellular and molecular biology, 2nd ed., vol. 2. ASM Press, Washington, D.C.
-
- Blattner, F. R., G. Plunkett III, C. A. Bloch, N. T. Perna, V. Burland, M. Riley, J. Collado-Vides, J. D. Glasner, C. K. Rode, G. F. Mayhew, J. Gregor, N. W. Davis, H. A. Kirkpatrick, M. A. Goeden, D. J. Rose, B. Mao, and Y. Shao. 1997. The complete genome sequence of Escherichia coli K-12. Science 277:1453–1474. - PubMed
-
- Bolivar, F., R. L. Rodriquez, P. J. Greene, M. C. Betlach, H. L. Heyneker, H. W. Boyer, J. H. Crosa, and S. Falkow. 1977. Construction and characterization of new cloning vehicles. II. A multipurpose cloning system. Gene 2:95–113. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
