

Ras and TGF[beta] cooperatively regulate epithelial cell plasticity and metastasis: dissection of Ras signaling pathways
- PMID: 11790801
- PMCID: PMC2199233
- DOI: 10.1083/jcb.200109037
Ras and TGF[beta] cooperatively regulate epithelial cell plasticity and metastasis: dissection of Ras signaling pathways
Abstract
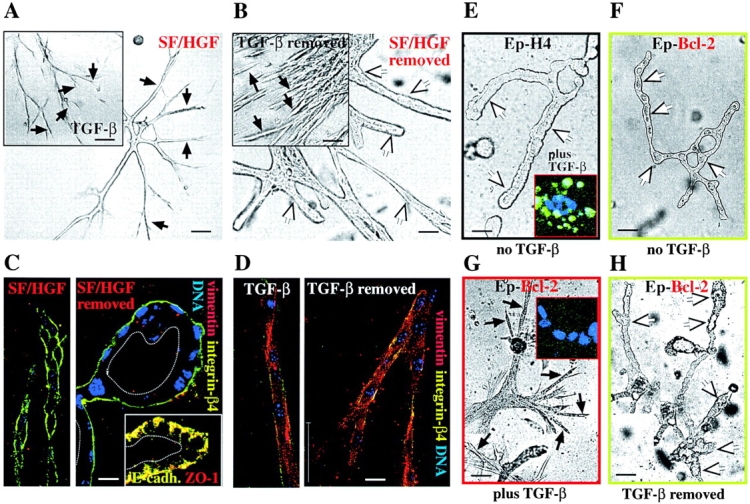
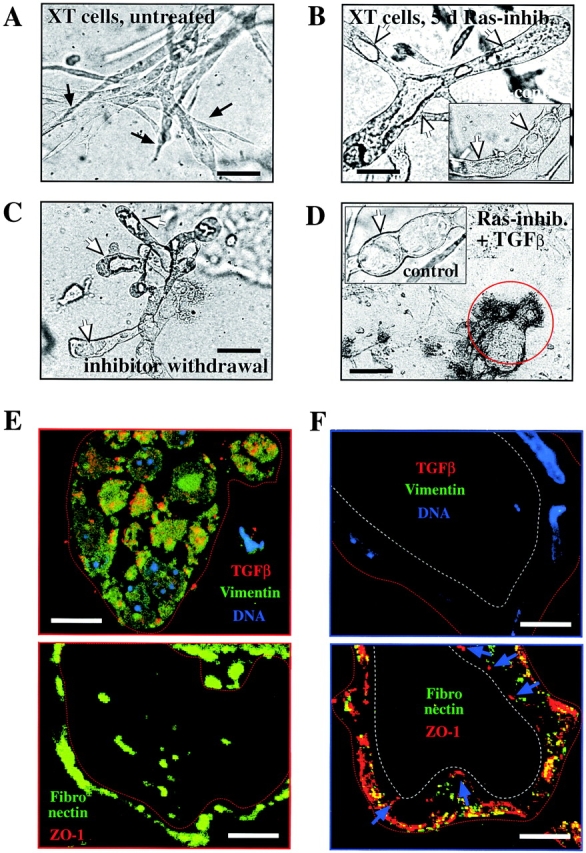
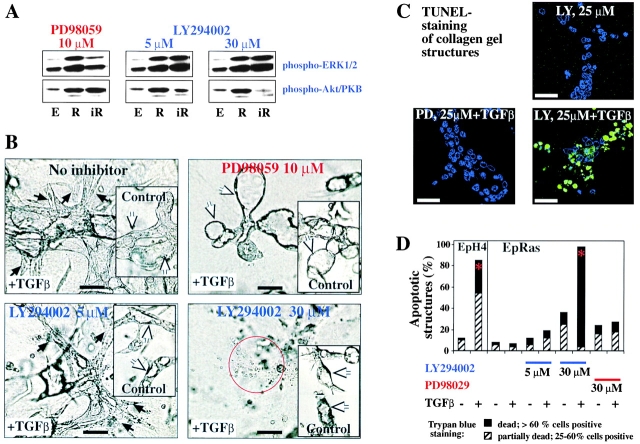
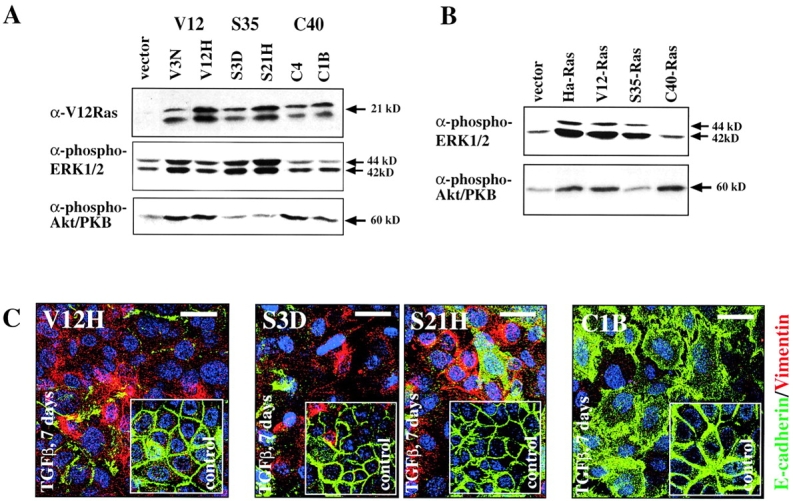
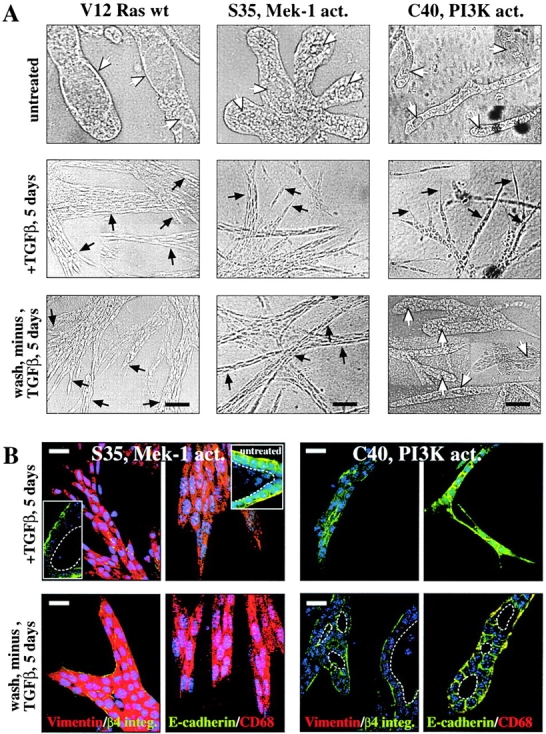
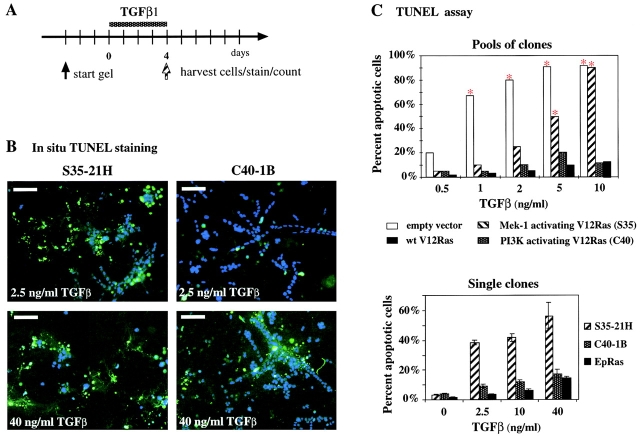
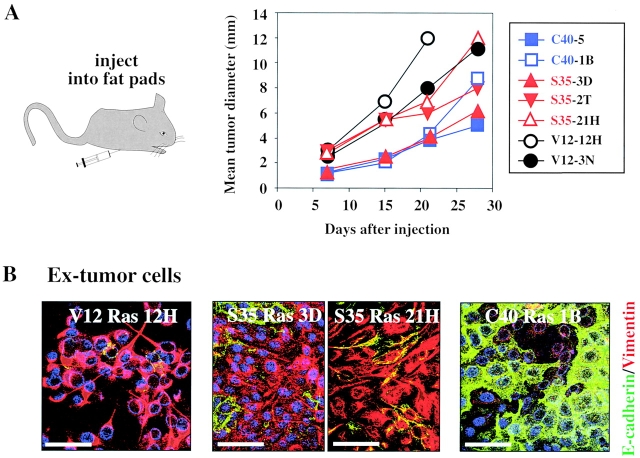
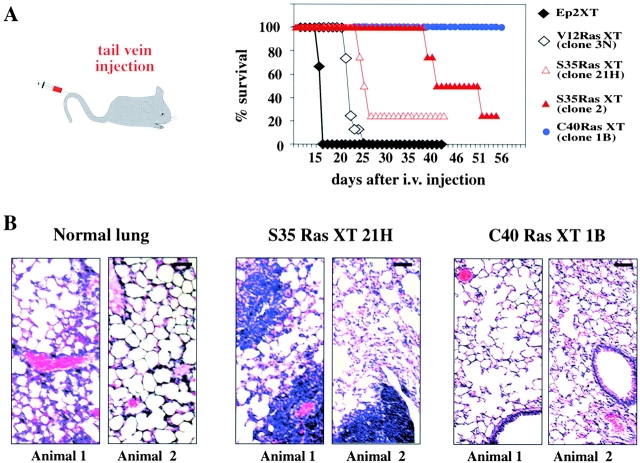
Multistep carcinogenesis involves more than six discrete events also important in normal development and cell behavior. Of these, local invasion and metastasis cause most cancer deaths but are the least well understood molecularly. We employed a combined in vitro/in vivo carcinogenesis model, that is, polarized Ha-Ras-transformed mammary epithelial cells (EpRas), to dissect the role of Ras downstream signaling pathways in epithelial cell plasticity, tumorigenesis, and metastasis. Ha-Ras cooperates with transforming growth factor beta (TGFbeta) to cause epithelial mesenchymal transition (EMT) characterized by spindle-like cell morphology, loss of epithelial markers, and induction of mesenchymal markers. EMT requires continuous TGFbeta receptor (TGFbeta-R) and oncogenic Ras signaling and is stabilized by autocrine TGFbeta production. In contrast, fibroblast growth factors, hepatocyte growth factor/scatter factor, or TGFbeta alone induce scattering, a spindle-like cell phenotype fully reversible after factor withdrawal, which does not involve sustained marker changes. Using specific inhibitors and effector-specific Ras mutants, we show that a hyperactive Raf/mitogen-activated protein kinase (MAPK) is required for EMT, whereas activation of phosphatidylinositol 3-kinase (PI3K) causes scattering and protects from TGFbeta-induced apoptosis. Hyperactivation of the PI3K pathway or the Raf/MAPK pathway are sufficient for tumorigenesis, whereas EMT in vivo and metastasis required a hyperactive Raf/MAPK pathway. Thus, EMT seems to be a close in vitro correlate of metastasis, both requiring synergism between TGFbeta-R and Raf/MAPK signaling.
Figures
References
-
- Bakin, A.V., A.K. Tomlinson, N.A. Bhowmick, H.A. Moses, and C.L. Arteaga. 2000. Phosphatidylinositol 3-kinase function is required for transforming growth factor β-mediated epithelial to mesenchymal transition and cell migration. J. Biol. Chem. 275:36803–36810. - PubMed
-
- Bergers, G., and L.M. Coussens. 2000. Extrinsic regulators of epithelial tumor progression: metalloproteinases. Curr. Opin. Genet. Dev. 10:120–127. - PubMed
-
- Boyer, B., A.M. Valles, and J.P. Thiery. 1996. Model systems of epithelium-mesenchyme transitions. Acta Anat. 156:227–239. - PubMed
-
- Boyer, B., A.M. Valles, and N. Edme. 2000. Induction and regulation of epithelial-mesenchymal transitions. Biochem. Pharmacol. 60:1091–1099. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
