

doi: 10.1086/338915.
Epub 2001 Dec 26.
2001 William Allan Award Address. From Down syndrome to the "human" in "human genetics"
Affiliations
- PMID: 11791206
- PMCID: PMC384910
- DOI: 10.1086/338915
Item in Clipboard
2001 William Allan Award Address. From Down syndrome to the "human" in "human genetics"
Am J Hum Genet.
2002 Feb.
No abstract available
Figures
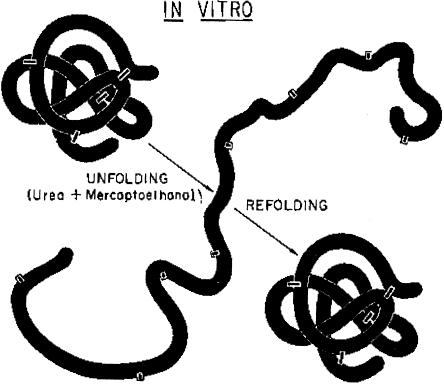
The experimental denaturation (UNFOLDING) and renaturation (REFOLDING) of a disulfide bond–containing protein, to demonstrate that the three-dimensional structure of the protein is determined by its primary amino acid sequence. Reprinted, by permission, from Epstein et al. (1963). (© 1963 by Cold Spring Harbor Laboratory Press.)
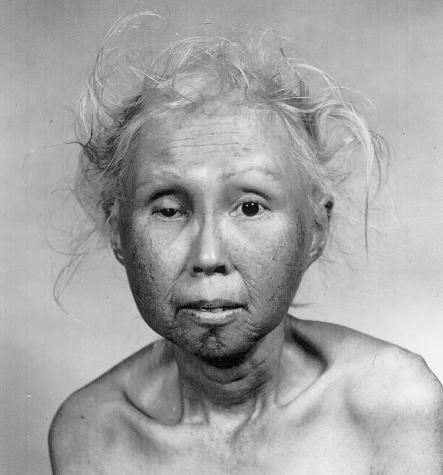
Forty-eight-year-old woman with Werner syndrome, an autosomal recessive disorder caused by a helicase mutation that produces a caricature of aging. Reprinted, by permission, from Epstein et al. (1966). (© 1966 by Lippincott Williams & Wilkins.)
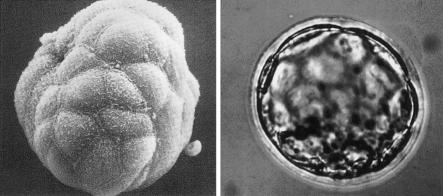
Scanning and transmission micrographs of a preimplantation mouse embryo at the blastocysts stage. Reprinted, by permission, from Calarco and Epstein (1973).

A trisomy 17 ↔ diploid chimera generated by preimplantation embryo aggregation, by the method developed by Beatrice Mintz (1972). Reprinted, by permission, from Epstein et al. (1982b). (© 1982 by the National Academy of Sciences, U.S.A.)
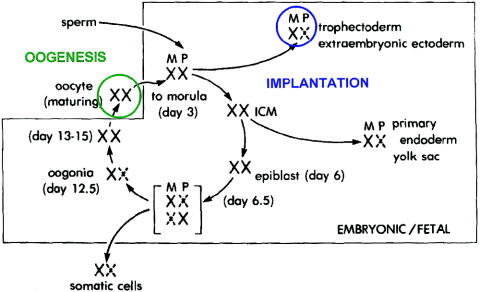
The life cycle of the X chromosome, showing that both X chromosomes are active during oogenesis and early embryonic development, with X-inactivation first occurring at the time of implantation. Reprinted, by permission, from Epstein (1981). (© 1981 by Lippincott Williams & Wilkins.)
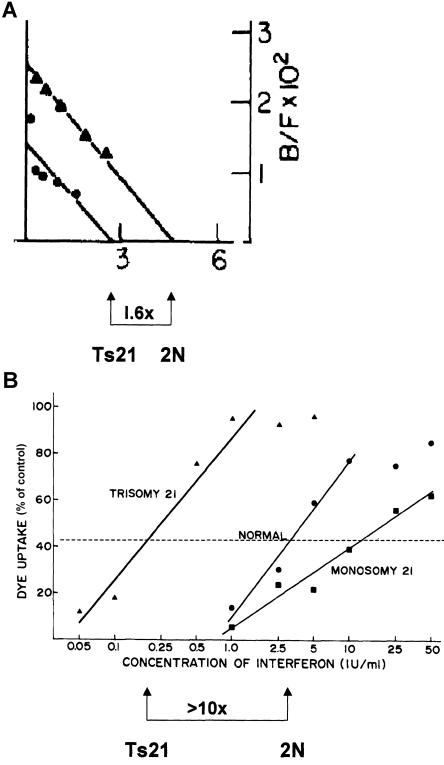
Interferon-α binding and antiviral sensitivity, in trisomy 21 cells. A, The binding of interferon-α to the interferon-α receptor in matched trisomy 21 and diploid fibroblasts. In the experiment shown, the trisomic cells bound 1.6 times as much interferon as do the diploid cells, which is very close to the theoretical value of 1.5 that is expected from the increase in copy number of the receptor gene from 2 to 3. Reprinted, by permission, from Epstein et al. (1982a). (© 1982 by Academic Press.) B, Increased sensitivity of trisomy 21 fibroblasts to the antiviral effect of interferon-α. In the experiment shown, sensitivity was increased by greater than 10-fold, but in several experiments the median increase was close to 6-fold. Reprinted, by permission, from Weil et al. (1980). (© 1980 by Mary Ann Liebert, Inc.)
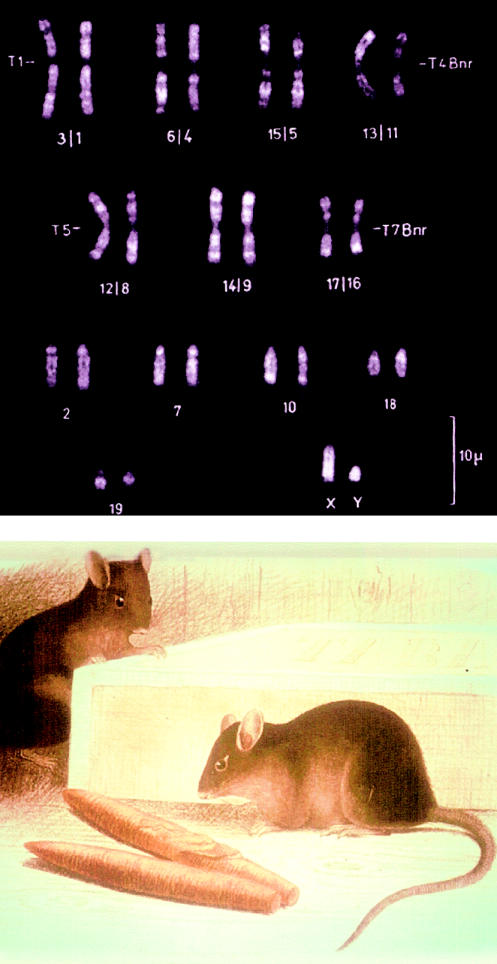
Fluorescent-banded karyotype (top) and somewhat fanciful picture of M. poschiavinus, the tobacco mouse (bottom). These pictures were obtained from the late Prof. Alfred Gropp, of Lübeck, Germany.
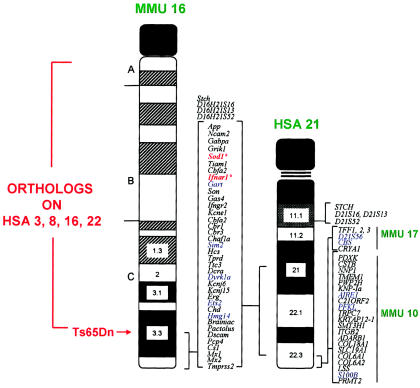
Syntenic region in common between human chromosome 21 (HSA 21) and mouse chromosome 16 (MMU 16) that forms the basis for the development of the trisomy 16 mouse models of Down syndrome. The other mouse chromosome regions orthologous to human chromosome 21 and other human chromosome regions orthologous to mouse chromosome 16 are shown, as is the location of the Ts65Dn breakpoint above App. The first two human chromosome 21 loci mapped to mouse chromosome 16 by Cox et al. (1980)—SOD1 and IFNAR—are marked with by asterisks (*). This figure is a modification of comparative maps kindly provided by Roger Reeves and published in Richtsmeier et al. (2000). Reprinted by permission. (© 2000 by Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc.)

Components of Down syndrome phenotype present in persons with segmental trisomy 21. The regions of trisomy in JS and KJ, indicated by the vertical bars, are quite similar to that present in the Ts65Dn mouse. Modified and reprinted, by permission, from Korenberg et al. (1994). (© 1994 by the National Academy of Sciences, U.S.A.)
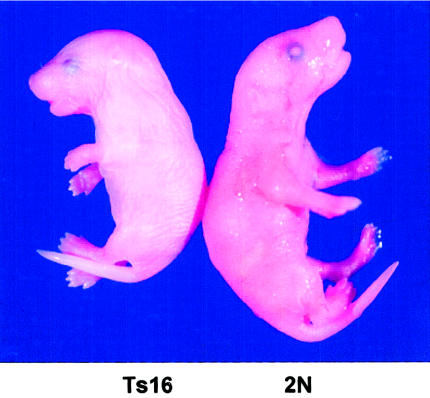
Mouse fetus with trisomy 16 (Ts16; left) and diploid littermate (2N; right). The small size, short snout, open eye lids, and thick neck (the residual of midgestational edema) caused by trisomy 16 are visible. Picture provided by David Holtzman (Washington University, St. Louis).
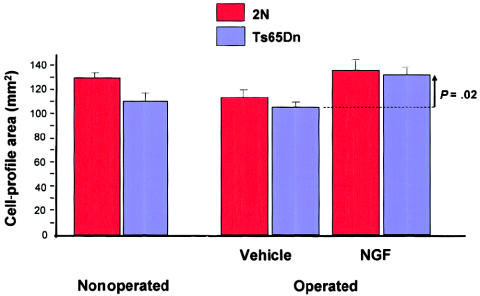
Restoration of size of basal-forebrain cholinergic neurons by infusion of nerve growth factor in aged Ts65n mice. The decrease in cell-profile area in the nonoperated Ts65Dn mice is significant at P=.009 , and the increase in cell-profile area in treated Ts65Dn animals (vertical arrow) is significant at P=.02 . Modified and reprinted, by permission, from Cooper et al. (2001). (© 2001 by the National Academy of Sciences, U.S.A.)
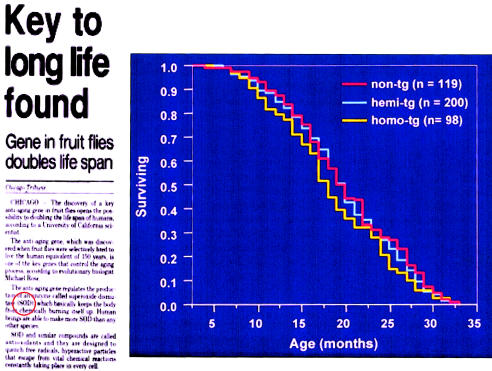
Association between CuZn SOD activity and life span. Although an association between elevated CuZn SOD activity and prolongation of life in Drosophila has been observed, transgenic mice with threefold (hemi-Tg) and fivefold (homo-Tg) increased CuZn SOD activity do not have an increased life span or an increased mean survival. Reprinted, by permission, from the Chicago Tribune and Huang et al. (2000). (© 2000 by the Gerontological Society of America.)
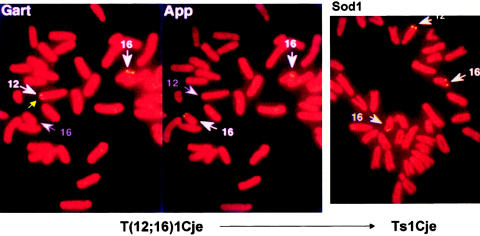
Reciprocal translocation and the rise of Ts1Cje. The identification of the T1Cje translocation by FISH. The left and middle panels identify the locations of Gart and App, respectively. Whereas the two App signals are located on chromosome 16, one of the more distal Gart signals is located on chromosome 12, indicating the presence of a reciprocal 12;16 translocation with a breakpoint between the two markers. In the trisomic Ts65Dn progeny of the balanced translocation carriers (right panel), there are three Sod1 signals, two on chromosome 16 and one on chromosome 12. The gene represented by the Sod1 signal on chromosome 12, which is at or just distal to the breakpoint, has been knocked out by the same gene-targeting event that apparently caused the translocation. The figures, prepared as described by Sago et al. (1998), were provided by Dr. Haru Sago.
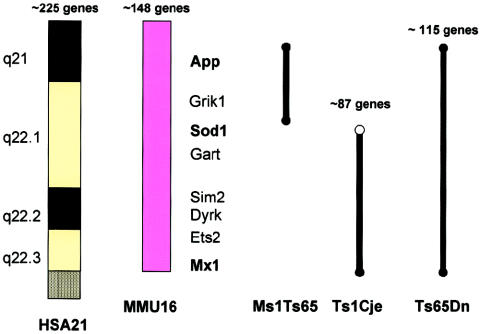
The three currently existing segmental trisomies for mouse chromosome 16. The numbers of genes in each trisomic segment represent approximations, subject to change, that were provided by Dr. Roger Reeves. Reprinted, by permission, from Epstein (2001). (© 2001 by the McGraw-Hill Companies.)
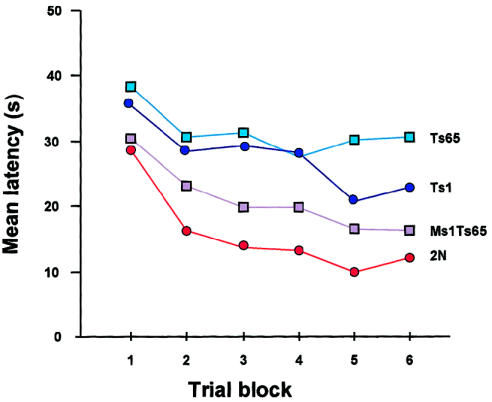
Latency (time to reach platform) in reverse hidden-platform test in the Morris water maze. The best performance is by 2N animals, and the poorest performance is by Ts65Dn. The performance by Ts1Cje is somewhat better than that by Ts65Dn, whereas that by Ts1Cje is better than that by the other two segmental trisomics but still not as good as that by the 2N mice. All mice were generated from the same Ts65Dn × T1Cje cross. Modified and reprinted, by permission, from Sago et al. (2000). (© 2000 by the International Pediatric Research Foundation, Inc.)

Comparison between skulls of Ts1Cje (top) and Ts65Dn (bottom) mice, demonstrating that there is a similar shortening of distances between landmarks (numerals) relative to diploid controls in the two segmental trisomics. Modified and reprinted, by permission, from Reeves et al. (2001) (© 2001 by Elsevier Science) and Richtsmeier et al. (in press) (© 2001 by Wiley-Liss, Inc., a subsidiary of John Wiley & Sons, Inc.).
The challenge facing human geneticists, as articulated by Epstein (1997).
The challenge facing human geneticists, with consideration of the human being entered into the balance.
References
-
- Akeson EC, Lambert JP, Narayanswami S, Gardiner K, Bechtel LJ, Davisson MT (2001) Ts65Dn: localization of the translocation breakpoint and trisomic gene content in a mouse model for Down syndrome. Cytogenet Cell Genet 93:270–276 - PubMed
-
- Anfinsen CB (1973) Principles that govern the folding of protein chains. Science 181:223–230 - PubMed
-
- Berger CN, Epstein CJ (1989) Delayed thymocyte maturation in the trisomy 16 mouse fetus. J Immunol 143:389–396 - PubMed
-
- Brinster RL (1965) Lactate dehydrogenase activity in the preimplanted mouse embryo. Biochim Biophys Acta 110:439–441 - PubMed
Publication types
MeSH terms
Personal name as subject
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
