

Self-assembly properties of a model RING domain
- PMID: 11792829
- PMCID: PMC117363
- DOI: 10.1073/pnas.012317299
Self-assembly properties of a model RING domain
Abstract
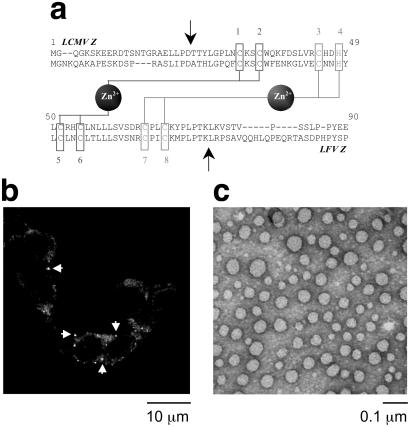
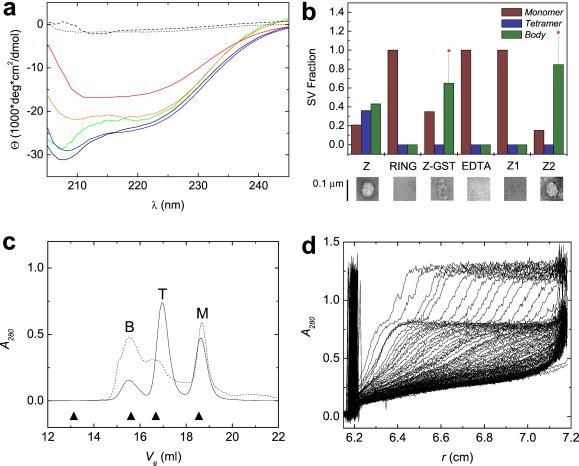
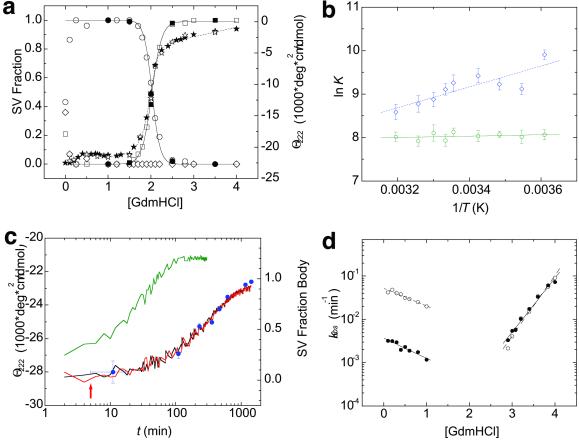
RING domains act in a variety of essential cellular processes but have no general function ascribed to them. Here, we observe that purified arenaviral protein Z, constituted almost entirely by its RING domain, self-assembles in vitro into spherical structures that resemble functional bodies formed by Z in infected cells. By using a variety of biophysical methods we provide a thermodynamic and kinetic framework for the RING-dependent self-assembly of Z. Assembly appears coupled to substantial conformational reorganization and changes in zinc coordination of site II of the RING. Thus, the rate-limiting nature of conformational reorganization observed in the folding of monomeric proteins can also apply to the assembly of macromolecular scaffolds. These studies describe a unique mechanism of nonfibrillar homogeneous self-assembly and suggest a general function of RINGs in the formation of macromolecular scaffolds that are positioned to integrate biochemical processes in cells.
Figures
Similar articles
-
Control of biochemical reactions through supramolecular RING domain self-assembly.Proc Natl Acad Sci U S A. 2002 Nov 26;99(24):15404-9. doi: 10.1073/pnas.202608799. Epub 2002 Nov 18. Proc Natl Acad Sci U S A. 2002. PMID: 12438698 Free PMC article.
-
The small RING finger protein Z drives arenavirus budding: implications for antiviral strategies.Proc Natl Acad Sci U S A. 2003 Oct 28;100(22):12978-83. doi: 10.1073/pnas.2133782100. Epub 2003 Oct 16. Proc Natl Acad Sci U S A. 2003. PMID: 14563923 Free PMC article.
-
Physical mechanisms and biological significance of supramolecular protein self-assembly.Curr Protein Pept Sci. 2004 Apr;5(2):125-34. doi: 10.2174/1389203043486856. Curr Protein Pept Sci. 2004. PMID: 15078223 Review.
-
Construction of macromolecular assemblages in eukaryotic processes and their role in human disease: linking RINGs together.Curr Protein Pept Sci. 2000 Jul;1(1):49-73. doi: 10.2174/1389203003381478. Curr Protein Pept Sci. 2000. PMID: 12369920
-
Review: conformation and folding of novel beta-structural elements in viral fiber proteins: the triple beta-spiral and triple beta-helix.J Struct Biol. 2002 Jan-Feb;137(1-2):236-47. doi: 10.1006/jsbi.2002.4447. J Struct Biol. 2002. PMID: 12064949 Review.
Cited by
-
The Mdm2 RING domain C-terminus is required for supramolecular assembly and ubiquitin ligase activity.EMBO J. 2007 Jan 10;26(1):90-101. doi: 10.1038/sj.emboj.7601465. Epub 2006 Dec 14. EMBO J. 2007. PMID: 17170710 Free PMC article.
-
Regulation of zinc homeostasis by inducible NO synthase-derived NO: nuclear metallothionein translocation and intranuclear Zn2+ release.Proc Natl Acad Sci U S A. 2003 Nov 25;100(24):13952-7. doi: 10.1073/pnas.2335190100. Epub 2003 Nov 14. Proc Natl Acad Sci U S A. 2003. PMID: 14617770 Free PMC article.
-
Multiple Weak Linear Motifs Enhance Recruitment and Processivity in SPOP-Mediated Substrate Ubiquitination.J Mol Biol. 2016 Mar 27;428(6):1256-1271. doi: 10.1016/j.jmb.2015.10.002. Epub 2015 Oct 22. J Mol Biol. 2016. PMID: 26475525 Free PMC article.
-
Arenavirus Z protein as an antiviral target: virus inactivation and protein oligomerization by zinc finger-reactive compounds.J Gen Virol. 2006 May;87(Pt 5):1217-1228. doi: 10.1099/vir.0.81667-0. J Gen Virol. 2006. PMID: 16603524 Free PMC article.
-
eIF4E orchestrates mRNA processing, RNA export and translation to modify specific protein production.Nucleus. 2024 Dec;15(1):2360196. doi: 10.1080/19491034.2024.2360196. Epub 2024 Jun 16. Nucleus. 2024. PMID: 38880976 Free PMC article. Review.
References
-
- Kentsis A, Borden K L B. Curr Protein Peptide Sci. 2000;1:49–73. - PubMed
-
- Peng H, Begg G E, Schultz D C, Friedman J R, Jensen D E, Speicher D W, Rauscher F J., III J Mol Biol. 2000;295:1139–1162. - PubMed
-
- Kentsis A, Dwyer E C, Perez J M, Sharma M, Chen A, Pan Z Q, Borden K L. J Mol Biol. 2001;312:609–623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
