

Requirement of an intact microtubule cytoskeleton for aggregation and inclusion body formation by a mutant huntingtin fragment
- PMID: 11792857
- PMCID: PMC117373
- DOI: 10.1073/pnas.022628699
Requirement of an intact microtubule cytoskeleton for aggregation and inclusion body formation by a mutant huntingtin fragment
Abstract
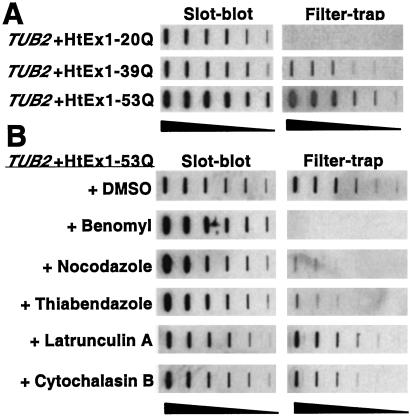
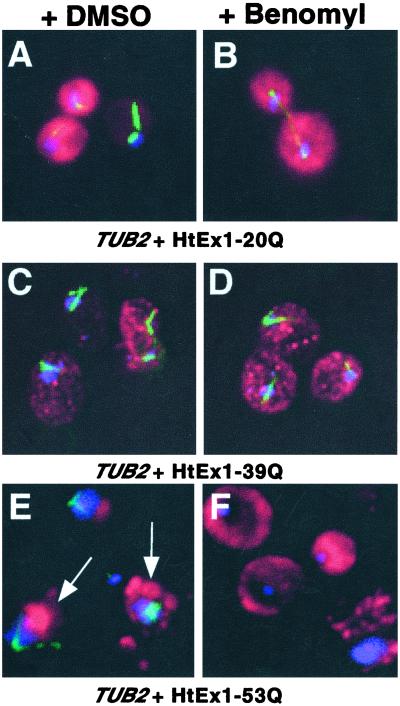
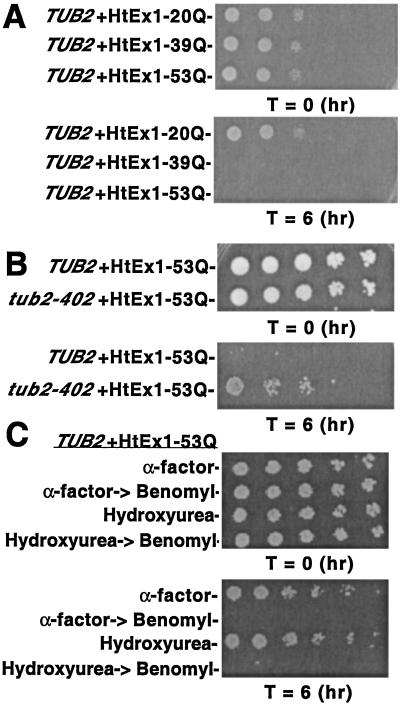
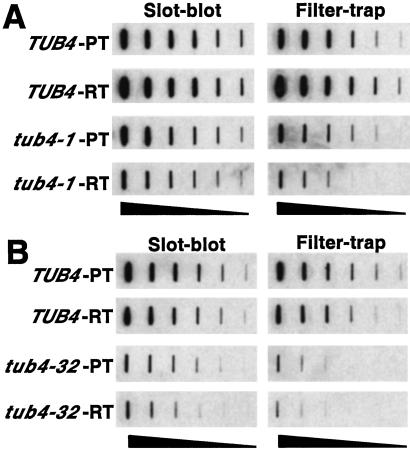
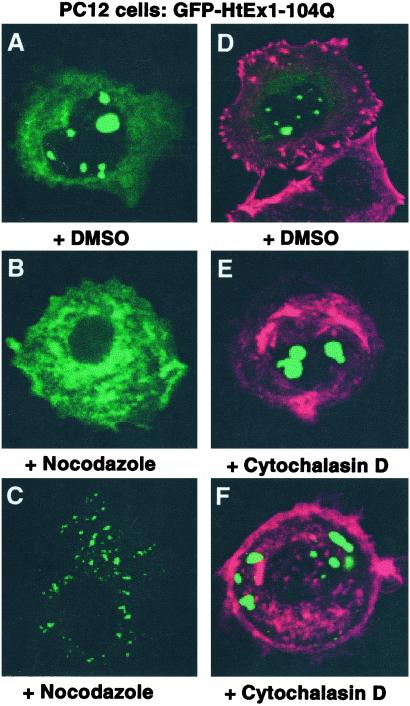
Huntington's disease is caused by the expansion of CAG repeats coding for a polyglutamine tract in the huntingtin protein. The major pathological feature found in Huntington's disease neurons is the presence of detergent-insoluble ubiquitinated inclusion bodies composed of the huntingtin protein. However, the mechanisms that underlie inclusion body formation, and the precise relationship between inclusion bodies and events that initiate toxicity, remain unclear. Here, we analyzed the effects of drugs or genetic mutations that disrupt the microtubule cytoskeleton in a Saccharomyces cerevisiae model of the aggregation of an amino-terminal polyglutamine-containing fragment of huntingtin exon 1 (HtEx1). Treatment of yeast with drugs that disrupt microtubules resulted in less than 2% of the detergent-insoluble HtEx1 observed in mock-treated cells and prevented the formation of large juxtanuclear inclusion bodies. Disruption of microtubules also unmasked a potent glutamine length-dependent toxicity of HtEx1 under conditions where HtEx1 exists in an entirely detergent-soluble nonaggregated form. Results from the yeast model paralleled those from neuronal pheochromocytoma cells, where disruption of microtubules eliminated the formation of juxtanuclear and intranuclear inclusion bodies by HtEx1. Our results suggest that active transport along microtubules may be required for inclusion body formation by HtEx1 and that inclusion body formation may have evolved as a cellular mechanism to promote the sequestration or clearance of soluble species of HtEx1 that are otherwise toxic to cells.
Figures
References
-
- Zoghbi H Y, Orr H T. Annu Rev Neurosci. 2000;23:217–247. - PubMed
-
- Saudou F, Finkbeiner S, Devys D, Greenberg M E. Cell. 1998;95:55–66. - PubMed
-
- Huynh D P, Figueroa K, Hoang N, Pulst S M. Nat Genet. 2000;26:44–50. - PubMed
-
- Warrick J M, Chan H Y, Gray-Board G L, Chai Y, Paulson H L, Bonini N M. Nat Genet. 1999;23:425–428. - PubMed
-
- Kazemi-Esfarjani P, Benzer S. Science. 2000;287:1837–1840. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
