

Differential binding of host complement inhibitor factor H by Borrelia burgdorferi Erp surface proteins: a possible mechanism underlying the expansive host range of Lyme disease spirochetes
- PMID: 11796574
- PMCID: PMC127719
- DOI: 10.1128/IAI.70.2.491-497.2002
Differential binding of host complement inhibitor factor H by Borrelia burgdorferi Erp surface proteins: a possible mechanism underlying the expansive host range of Lyme disease spirochetes
Abstract
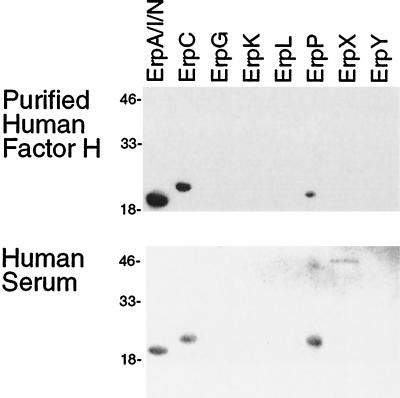
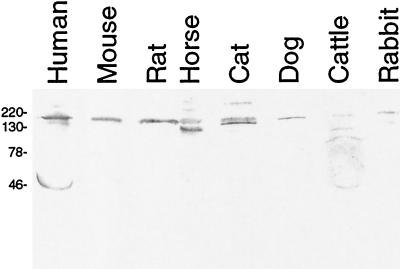
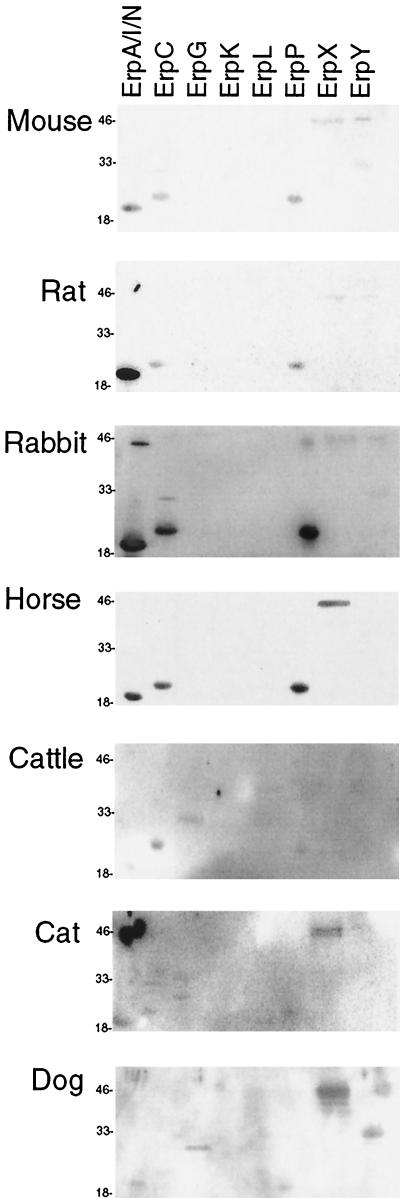
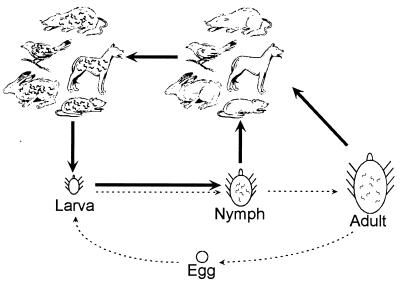
The Lyme disease spirochete, Borrelia burgdorferi, is capable of infecting a wide variety of vertebrates. This broad host range implies that B. burgdorferi possesses the ability to contravene the immune defenses of many potential hosts. B. burgdorferi produces multiple different Erp proteins on its outer membrane during mammalian infection. It was reported previously that one Erp protein can bind human factor H (J. Hellwage, T. Meri, T. Heikkilä, A. Alitalo, J. Panelius, P. Lahdenne, I. J. T. Seppälä, and S. Meri, J. Biol. Chem. 276:8427-8435, 2001). In this paper we report that the ability to bind the complement inhibitor factor H is a general characteristic of Erp proteins. Furthermore, each Erp protein exhibits different relative affinities for the complement inhibitors of various potential animal hosts. The data suggest that the presence of multiple Erp proteins on the surface can allow a single B. burgdorferi bacterium to resist complement-mediated killing in any of the wide range of potential hosts that it might infect. Thus, Erp proteins likely contribute to the persistence of B. burgdorferi in nature and to the ability of this bacterium to cause Lyme disease in humans and other animals.
Figures
Similar articles
-
Increased expression of Borrelia burgdorferi factor H-binding surface proteins during transmission from ticks to mice.Int J Med Microbiol. 2004 Apr;293 Suppl 37:120-5. doi: 10.1016/s1433-1128(04)80020-2. Int J Med Microbiol. 2004. PMID: 15146993
-
Complement resistance of Borrelia burgdorferi correlates with the expression of BbCRASP-1, a novel linear plasmid-encoded surface protein that interacts with human factor H and FHL-1 and is unrelated to Erp proteins.J Biol Chem. 2004 Jan 23;279(4):2421-9. doi: 10.1074/jbc.M308343200. Epub 2003 Nov 7. J Biol Chem. 2004. PMID: 14607842
-
Functional characterization of BbCRASP-2, a distinct outer membrane protein of Borrelia burgdorferi that binds host complement regulators factor H and FHL-1.Mol Microbiol. 2006 Sep;61(5):1220-36. doi: 10.1111/j.1365-2958.2006.05318.x. Mol Microbiol. 2006. PMID: 16925556
-
Lyme borreliosis spirochete Erp proteins, their known host ligands, and potential roles in mammalian infection.Int J Med Microbiol. 2008 Sep 1;298 Suppl 1(Suppl 1):257-67. doi: 10.1016/j.ijmm.2007.09.004. Epub 2008 Jan 11. Int J Med Microbiol. 2008. PMID: 18248770 Free PMC article. Review.
-
New Insights Into CRASP-Mediated Complement Evasion in the Lyme Disease Enzootic Cycle.Front Cell Infect Microbiol. 2020 Jan 30;10:1. doi: 10.3389/fcimb.2020.00001. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32083019 Free PMC article. Review.
Cited by
-
Erp and Rev Adhesins of the Lyme Disease Spirochete's Ubiquitous cp32 Prophages Assist the Bacterium during Vertebrate Infection.Infect Immun. 2023 Mar 15;91(3):e0025022. doi: 10.1128/iai.00250-22. Epub 2023 Feb 28. Infect Immun. 2023. PMID: 36853019 Free PMC article. Review.
-
Borrelia burgdorferi BmpA is a laminin-binding protein.Infect Immun. 2009 Nov;77(11):4940-6. doi: 10.1128/IAI.01420-08. Epub 2009 Aug 24. Infect Immun. 2009. PMID: 19703983 Free PMC article.
-
Analysis of the ability of spirochete species associated with relapsing fever, avian borreliosis, and epizootic bovine abortion to bind factor H and cleave c3b.J Clin Microbiol. 2003 Aug;41(8):3905-10. doi: 10.1128/JCM.41.8.3905-3910.2003. J Clin Microbiol. 2003. PMID: 12904415 Free PMC article.
-
Does host complement kill Borrelia burgdorferi within ticks?Infect Immun. 2003 Feb;71(2):822-9. doi: 10.1128/IAI.71.2.822-829.2003. Infect Immun. 2003. PMID: 12540562 Free PMC article.
-
Delineating the requirement for the Borrelia burgdorferi virulence factor OspC in the mammalian host.Infect Immun. 2006 Jun;74(6):3547-53. doi: 10.1128/IAI.00158-06. Infect Immun. 2006. PMID: 16714587 Free PMC article.
References
-
- Alexander, J. J., B. K. Hack, P. N. Cunningham, and R. J. Quigg. 2001. A protein with characteristics of factor H is present on rodent platelets and functions as the immune adherence receptor. J. Biol. Chem. 276:32129–32135. - PubMed
-
- Appel, M. J. G., S. Allan, R. H. Jacobson, T. L. Lauderdale, Y. F. Chang, S. J. Shin, J. W. Thomford, R. J. Todhunter, and B. A. Summers. 1993. Experimental Lyme disease in dogs produces arthritis and persistent infection. J. Infect. Dis. 167:651–664. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
