

Recurrent evolution of DNA-binding motifs in the Drosophila centromeric histone
- PMID: 11805302
- PMCID: PMC122211
- DOI: 10.1073/pnas.032664299
Recurrent evolution of DNA-binding motifs in the Drosophila centromeric histone
Abstract
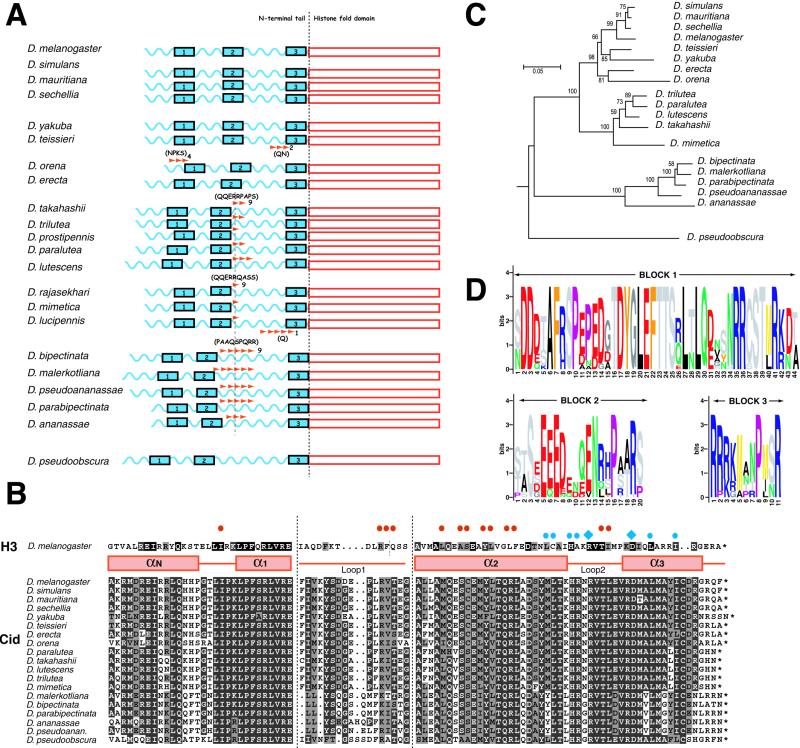
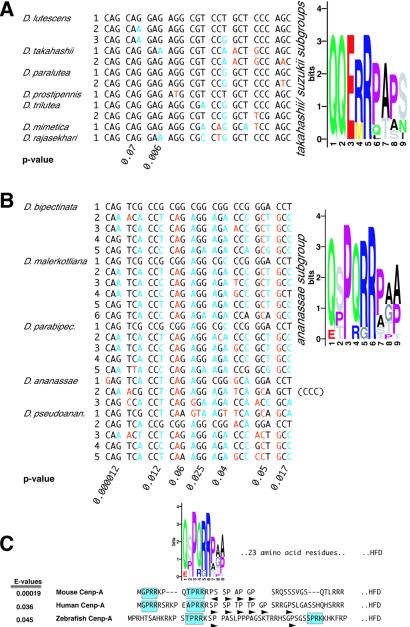
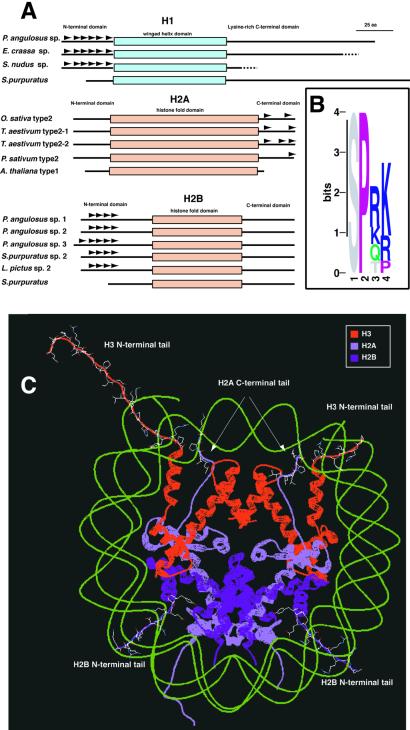
All eukaryotes contain centromere-specific histone H3 variants (CenH3s), which replace H3 in centromeric chromatin. We have previously documented the adaptive evolution of the Drosophila CenH3 (Cid) in comparisons of Drosophila melanogaster and Drosophila simulans, a divergence of approximately 2.5 million years. We have proposed that rapidly changing centromeric DNA may be driving CenH3's altered DNA-binding specificity. Here, we compare Cid sequences from a phylogenetically broader group of Drosophila species to suggest that Cid has been evolving adaptively for at least 25 million years. Our analysis also reveals conserved blocks not only in the histone-fold domain but also in the N-terminal tail. In several lineages, the N-terminal tail of Cid is characterized by subgroup-specific oligopeptide expansions. These expansions resemble minor groove DNA binding motifs found in various histone tails. Remarkably, similar oligopeptides are also found in N-terminal tails of human and mouse CenH3 (Cenp-A). The recurrent evolution of these motifs in CenH3 suggests a packaging function for the N-terminal tail, which results in a unique chromatin organization at the primary constriction, the cytological marker of centromeres.
Figures
Similar articles
-
Adaptive evolution of Cid, a centromere-specific histone in Drosophila.Genetics. 2001 Mar;157(3):1293-8. doi: 10.1093/genetics/157.3.1293. Genetics. 2001. PMID: 11238413 Free PMC article.
-
Adaptive evolution of the histone fold domain in centromeric histones.Mol Biol Evol. 2004 Sep;21(9):1712-8. doi: 10.1093/molbev/msh179. Epub 2004 Jun 2. Mol Biol Evol. 2004. PMID: 15175412
-
The rapidly evolving centromere-specific histone has stringent functional requirements in Arabidopsis thaliana.Genetics. 2010 Oct;186(2):461-71. doi: 10.1534/genetics.110.120337. Epub 2010 Jul 13. Genetics. 2010. PMID: 20628040 Free PMC article.
-
Centromeres Drive a Hard Bargain.Trends Genet. 2017 Feb;33(2):101-117. doi: 10.1016/j.tig.2016.12.001. Epub 2017 Jan 7. Trends Genet. 2017. PMID: 28069312 Free PMC article. Review.
-
Epigenetic specification of centromeres by CENP-A.Exp Cell Res. 2009 Nov 15;315(19):3233-41. doi: 10.1016/j.yexcr.2009.07.023. Epub 2009 Aug 3. Exp Cell Res. 2009. PMID: 19660450 Review.
Cited by
-
The role of DNA sequence in centromere formation.Genome Biol. 2003;4(5):214. doi: 10.1186/gb-2003-4-5-214. Epub 2003 Apr 29. Genome Biol. 2003. PMID: 12734002 Free PMC article. Review.
-
Evolution of sequence-specific anti-silencing systems in Arabidopsis.Nat Commun. 2017 Dec 18;8(1):2161. doi: 10.1038/s41467-017-02150-7. Nat Commun. 2017. PMID: 29255196 Free PMC article.
-
Characterization of CENH3 proteins and centromere-associated DNA sequences in diploid and allotetraploid Brassica species.Chromosoma. 2011 Aug;120(4):353-65. doi: 10.1007/s00412-011-0315-z. Epub 2011 Mar 11. Chromosoma. 2011. PMID: 21394438
-
The role of meiotic drive in hybrid male sterility.Philos Trans R Soc Lond B Biol Sci. 2010 Apr 27;365(1544):1265-72. doi: 10.1098/rstb.2009.0264. Philos Trans R Soc Lond B Biol Sci. 2010. PMID: 20308102 Free PMC article. Review.
-
Co-evolving CENP-A and CAL1 Domains Mediate Centromeric CENP-A Deposition across Drosophila Species.Dev Cell. 2016 Apr 18;37(2):136-47. doi: 10.1016/j.devcel.2016.03.021. Dev Cell. 2016. PMID: 27093083 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
