

Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm
- PMID: 11807075
- PMCID: PMC134825
- DOI: 10.1128/jb.184.4.1140-1154.2002
Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm
Abstract
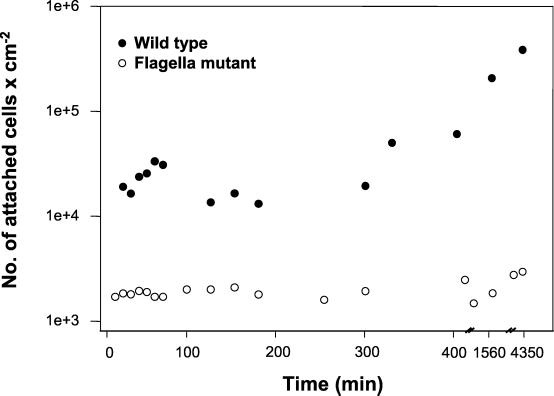
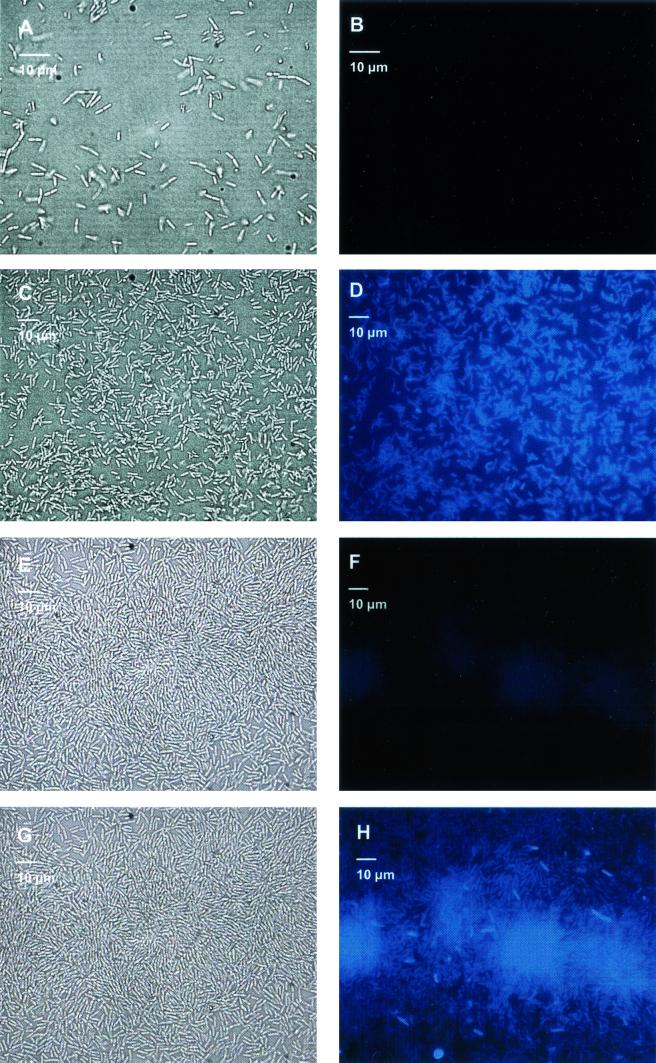
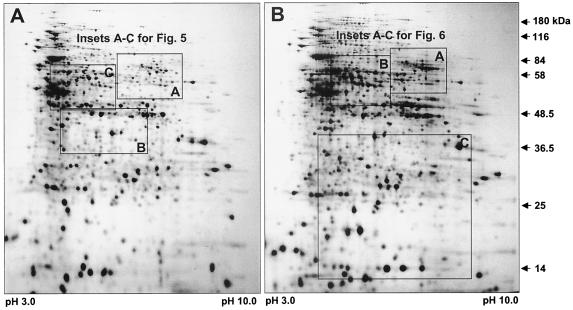
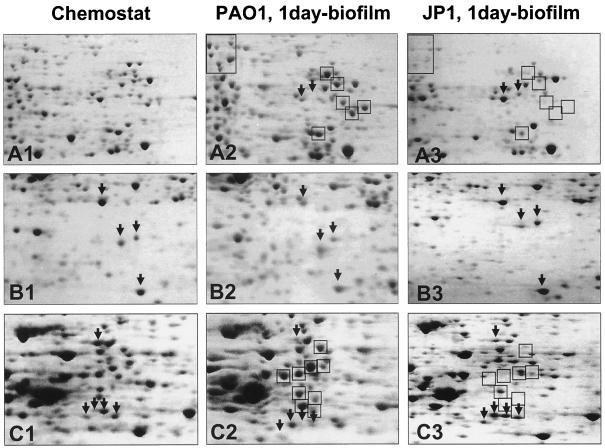
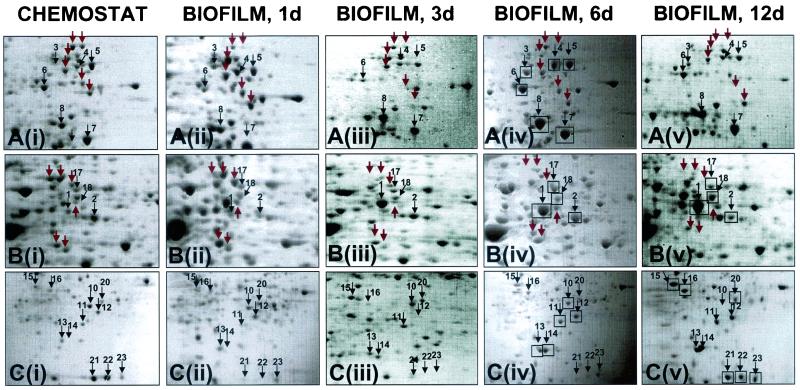
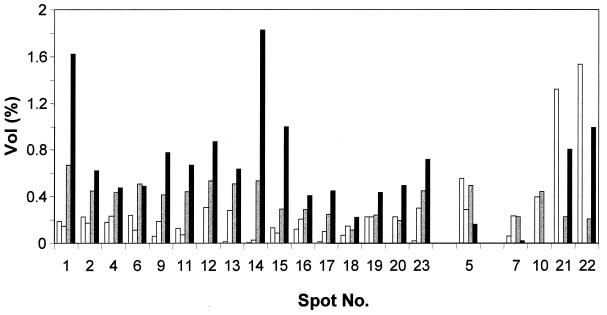
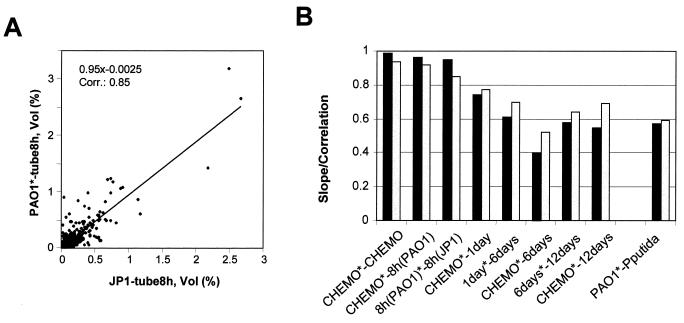
Complementary approaches were employed to characterize transitional episodes in Pseudomonas aeruginosa biofilm development using direct observation and whole-cell protein analysis. Microscopy and in situ reporter gene analysis were used to directly observe changes in biofilm physiology and to act as signposts to standardize protein collection for two-dimensional electrophoretic analysis and protein identification in chemostat and continuous-culture biofilm-grown populations. Using these approaches, we characterized five stages of biofilm development: (i) reversible attachment, (ii) irreversible attachment, (iii) maturation-1, (iv) maturation-2, and (v) dispersion. Biofilm cells were shown to change regulation of motility, alginate production, and quorum sensing during the process of development. The average difference in detectable protein regulation between each of the five stages of development was 35% (approximately 525 proteins). When planktonic cells were compared with maturation-2 stage biofilm cells, more than 800 proteins were shown to have a sixfold or greater change in expression level (over 50% of the proteome). This difference was higher than when planktonic P. aeruginosa were compared with planktonic cultures of Pseudomonas putida. Las quorum sensing was shown to play no role in early biofilm development but was important in later stages. Biofilm cells in the dispersion stage were more similar to planktonic bacteria than to maturation-2 stage bacteria. These results demonstrate that P. aeruginosa displays multiple phenotypes during biofilm development and that knowledge of stage-specific physiology may be important in detecting and controlling biofilm growth.
Figures

References
-
- Arai, H., T. Kodama, and Y. Igarashi. 1997. Cascade regulation of the two CRP/FNR-related transcriptional regulators (ANR and DNR) and the denitrification enzymes in Pseudomonas aeruginosa. Mol. Microbiol. 25:1141-1148. - PubMed
-
- Blum, H., H. Beier, and H. J. Gross. 1987. Improved silver staining of plant proteins, RNA and DNA in polyacrylamide gels. Electrophoresis 8:93-99.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
