

A new role for BiP: closing the aqueous translocon pore during protein integration into the ER membrane
- PMID: 11807091
- PMCID: PMC2199230
- DOI: 10.1083/jcb.200110074
A new role for BiP: closing the aqueous translocon pore during protein integration into the ER membrane
Abstract
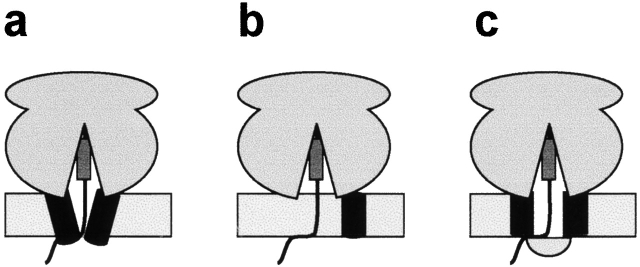
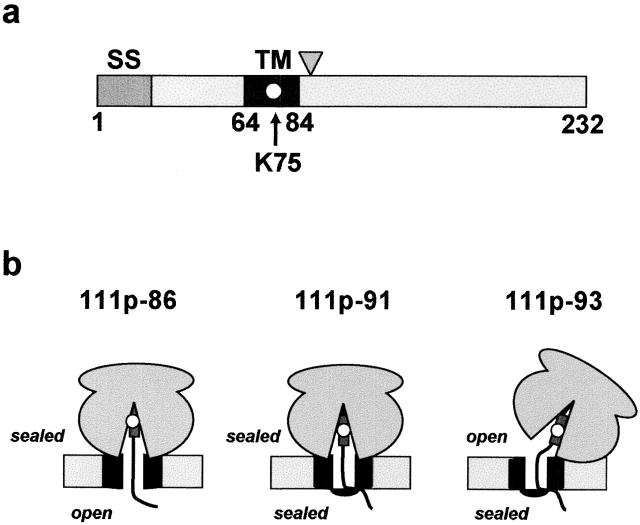
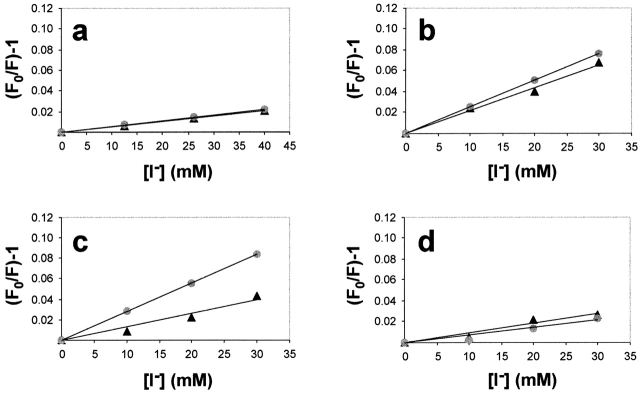
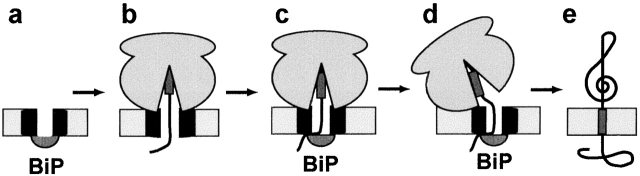
In mammalian cells, most membrane proteins are inserted cotranslationally into the ER membrane at sites termed translocons. Although each translocon forms an aqueous pore, the permeability barrier of the membrane is maintained during integration, even when the otherwise tight ribosome-translocon seal is opened to allow the cytoplasmic domain of a nascent protein to enter the cytosol. To identify the mechanism by which membrane integrity is preserved, nascent chain exposure to each side of the membrane was determined at different stages of integration by collisional quenching of a fluorescent probe in the nascent chain. Comparing integration intermediates prepared with intact, empty, or BiP-loaded microsomes revealed that the lumenal end of the translocon pore is closed by BiP in an ATP-dependent process before the opening of the cytoplasmic ribosome-translocon seal during integration. This BiP function is distinct from its previously identified role in closing ribosome-free, empty translocons because of the presence of the ribosome at the translocon and the nascent membrane protein that extends through the translocon pore and into the lumen during integration. Therefore, BiP is a key component in a sophisticated mechanism that selectively closes the lumenal end of some, but not all, translocons occupied by a nascent chain. By using collisional quenchers of different sizes, the large internal diameter of the ribosome-bound aqueous translocon pore was found to contract when BiP was required to seal the pore during integration. Therefore, closure of the pore involves substantial conformational changes in the translocon that are coupled to a complex sequence of structural rearrangements on both sides of the ER membrane involving the ribosome and BiP.
Figures
References
-
- Beckmann, R., D. Bubeck, R. Grassucci, P. Penczek, A. Verschoor, G. Blobel, and J. Frank. 1997. Alignment of conduits for the nascent polypeptide chain in the ribosome-Sec61 complex. Science. 278:2123–2126. - PubMed
-
- Beckmann, R., C.M.T. Spahn, N. Eswar, J. Helmers, P.A. Penczek, A. Sali, J. Frank, and G. Blobel. 2001. Architecture of the protein-conducting channel associated with the translating 80S ribosome. Cell. 107:361–372. - PubMed
-
- Bell, C.E., and D. Eisenberg. 1996. Crystal structure of diphtheria toxin bound to nicotinamide adenine dinucleotide. Biochemistry. 35:1137–1149. - PubMed
-
- Brodsky, J.L. 1996. Post-translational protein translocation: not all hsc70s are created equal. Trends Biochem. Sci. 21:122–126. - PubMed
-
- Brodsky, J.L., and A.A. McCracken. 1999. ER protein quality control and proteasome-mediated protein degradation. Semin. Cell Dev. Biol. 10:507–513. - PubMed
