

The integrin cytoplasmic domain-associated protein ICAP-1 binds and regulates Rho family GTPases during cell spreading
- PMID: 11807099
- PMCID: PMC2199222
- DOI: 10.1083/jcb.200108030
The integrin cytoplasmic domain-associated protein ICAP-1 binds and regulates Rho family GTPases during cell spreading
Abstract
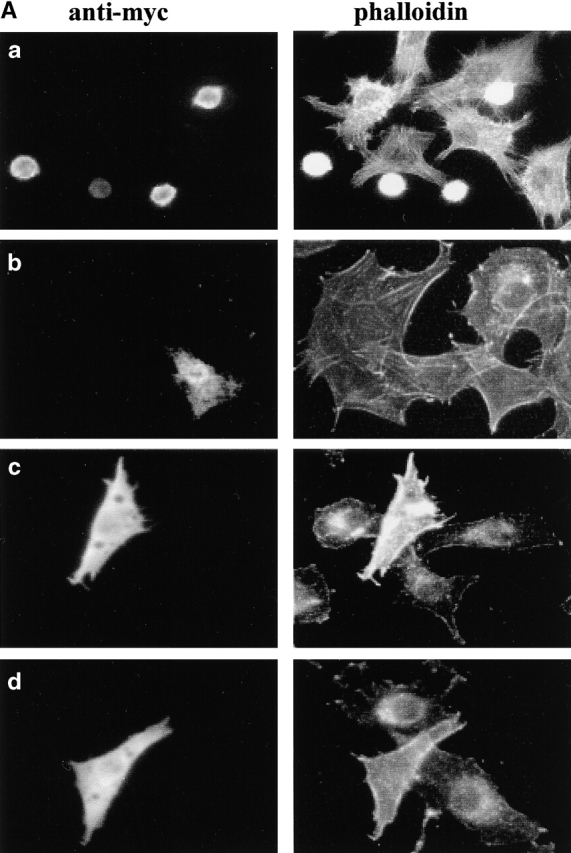
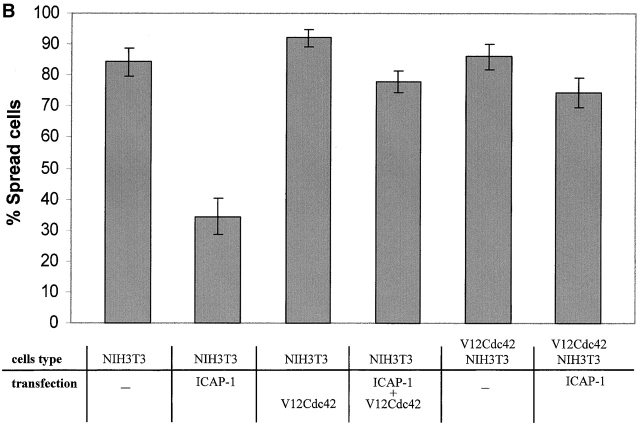
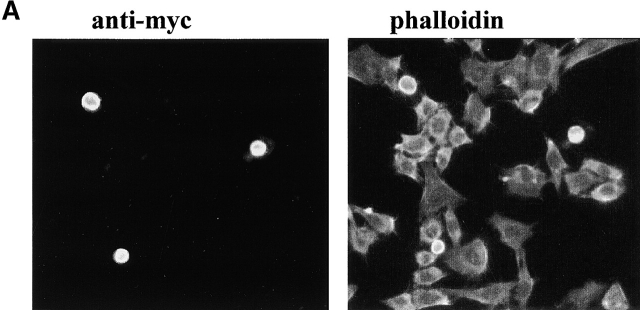
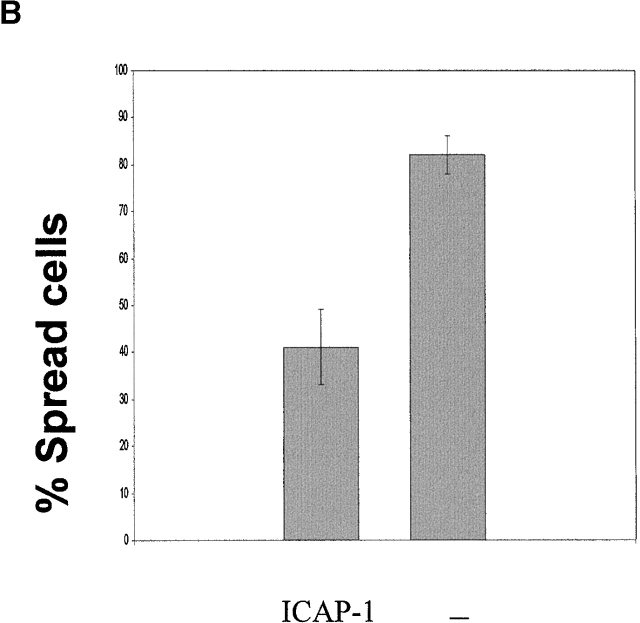
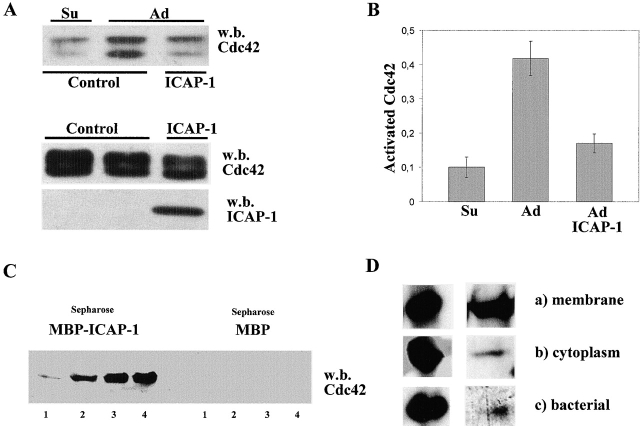
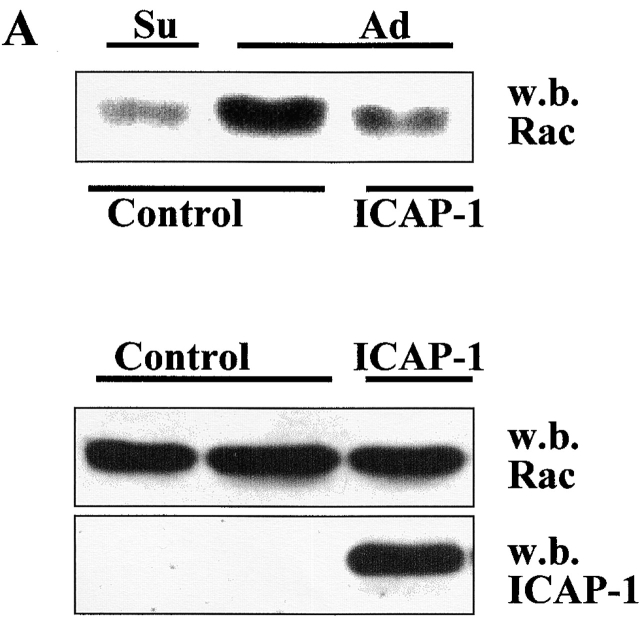
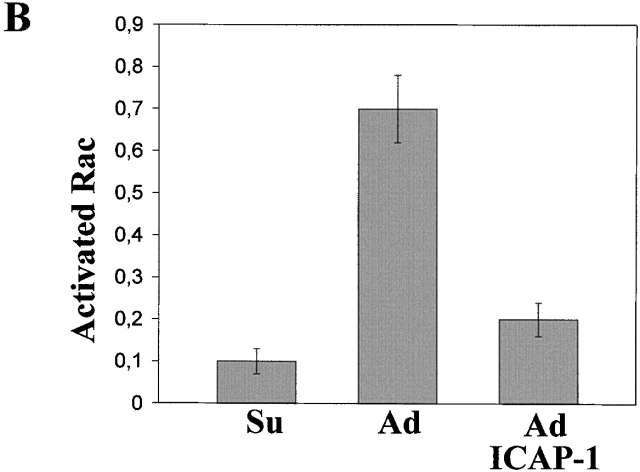
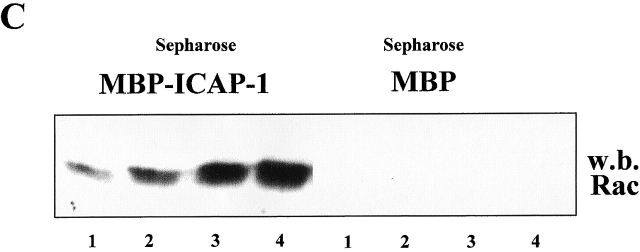
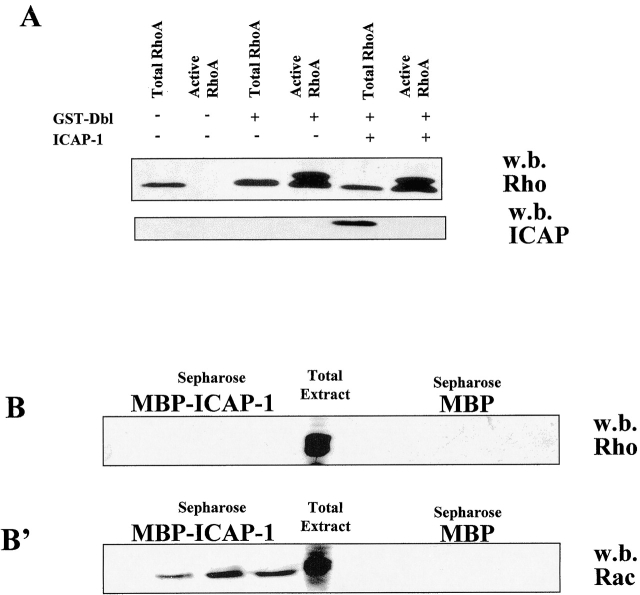
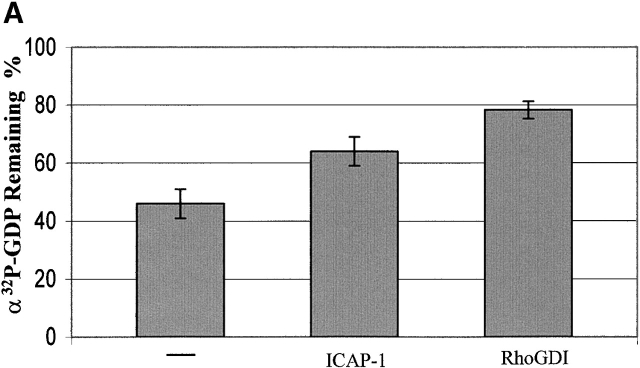
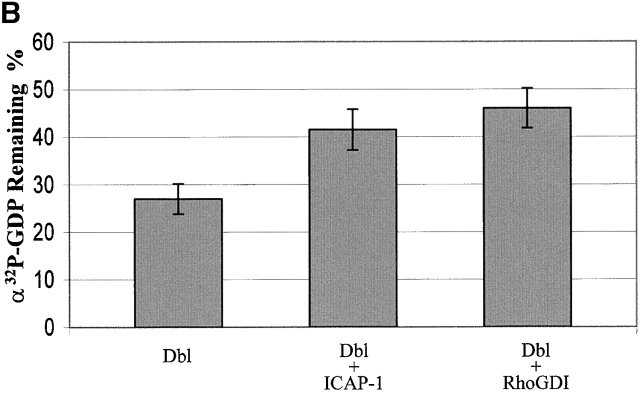
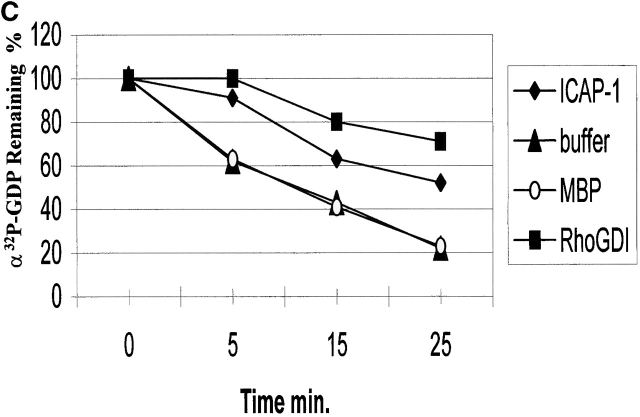
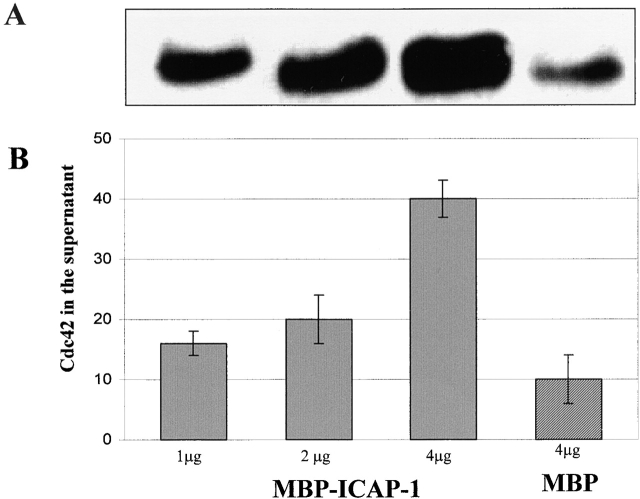
Using two-hybrid screening, we isolated the integrin cytoplasmic domain-associated protein (ICAP-1), an interactor for the COOH terminal region of the beta1A integrin cytoplasmic domain. To investigate the role of ICAP-1 in integrin-mediated adhesive function, we expressed the full-length molecule in NIH3T3 cells. ICAP-1 expression strongly prevents NIH3T3 cell spreading on extracellular matrix. This inhibition is transient and can be counteracted by coexpression of a constitutively activated mutant of Cdc42, suggesting that ICAP-1 acts upstream of this GTPase. In addition, we found that ICAP-1 binds both to Cdc42 and Rac1 in vitro, and its expression markedly inhibits activation of these GTPases during integrin-mediated cell adhesion to fibronectin as detected by PAK binding assay. In the attempt to define the molecular mechanism of this inhibition, we show that ICAP-1 reduces both the intrinsic and the exchange factor-induced dissociation of GDP from Cdc42; moreover, purified ICAP-1 displaces this GTPase from cellular membranes. Together, these data show for the first time that ICAP-1 regulates Rho family GTPases during integrin-mediated cell matrix adhesion, acting as guanine dissociation inhibitor.
Figures

References
-
- Arthur, W.T., L.A. Petch, and K. Burridge. 2000. Integrin engagement suppresses RhoA activity via a c-Src-dependent mechanism. Curr. Biol. 10:719–722. - PubMed
-
- Belkin, A.M., N.I. Zhidkova, F. Balzac, F. Altruda, D. Tomatis, A. Maier, G. Tarone, V.E. Koteliansky, and K. Burridge. 1996. Beta1D integrin displaces the beta 1A isoform in striated muscles: localization at junctional structures and signaling potential in nonmuscle cells. J. Cell Biol. 132:211–226. - PMC - PubMed
-
- Belkin, A.M., S.F. Retta, O.Y. Pletjushkina, F. Balzac, L. Silengo, R. Fassler, V.E. Koteliansky, K. Burridge, and G. Tarone. 1997. Muscle beta1D integrin reinforces the cytoskeleton-matrix link: modulation of integrin adhesive function by alternative splicing. J. Cell Biol. 139:1583–1595. - PMC - PubMed
-
- Bouvard, D., and M.R. Block. 1998. Calcium/calmodulin-dependent protein kinase II controls integrin alpha5 beta1-mediated cell adhesion through the integrin cytoplasmic domain associated protein-1alpha. Biochem. Biophys. Res. Commun. 252:46–50. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
