

Evidence for a dynamic-dominance hypothesis of handedness
- PMID: 11807578
- PMCID: PMC10710695
- DOI: 10.1007/s00221-001-0913-8
Evidence for a dynamic-dominance hypothesis of handedness
Abstract
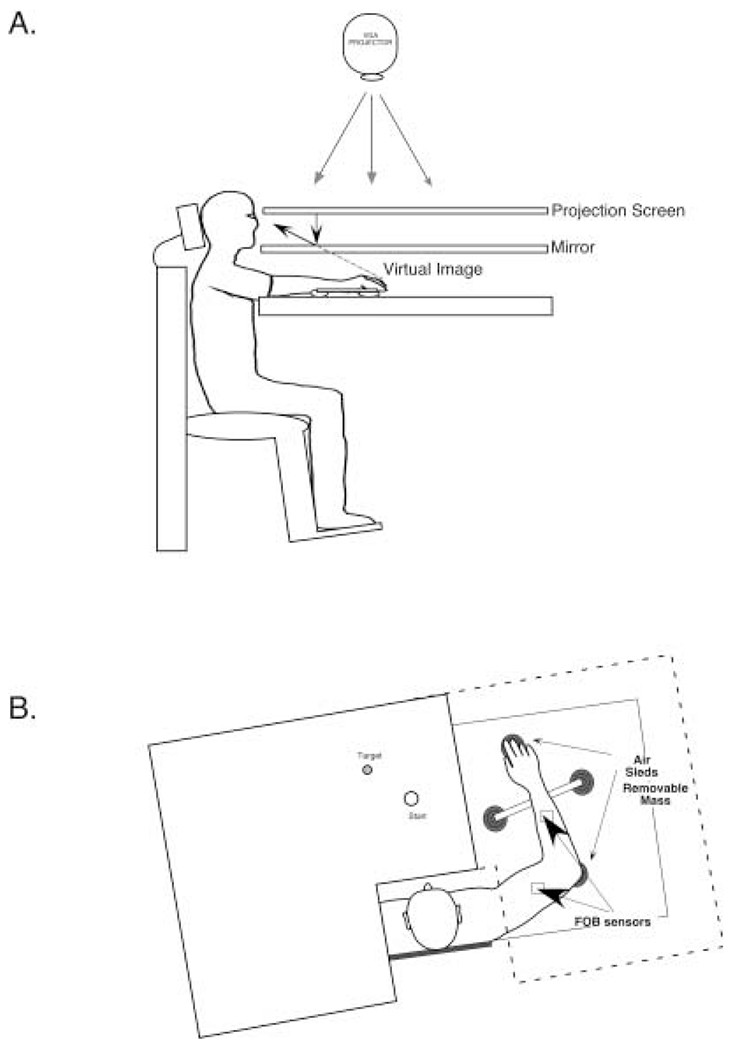
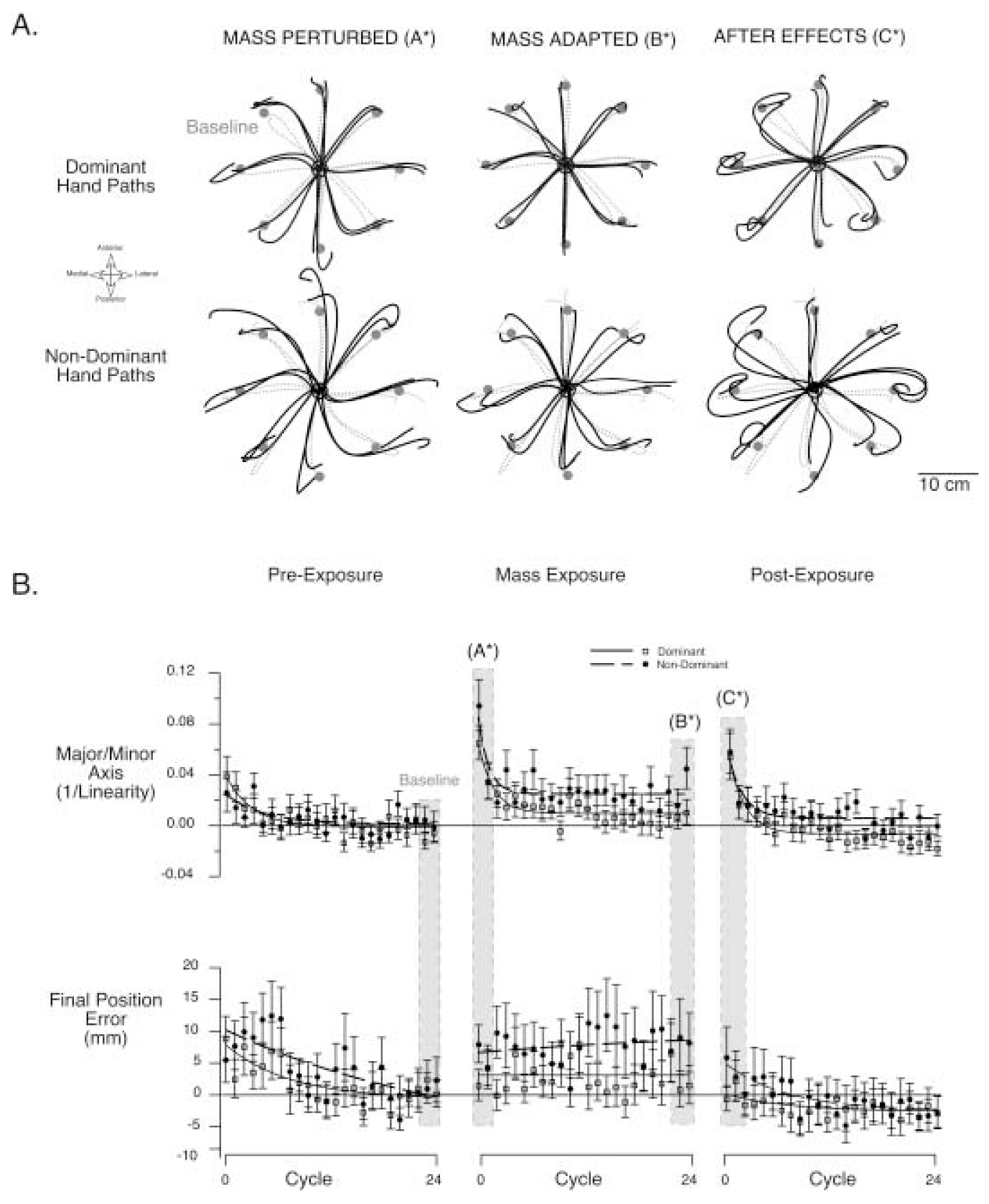
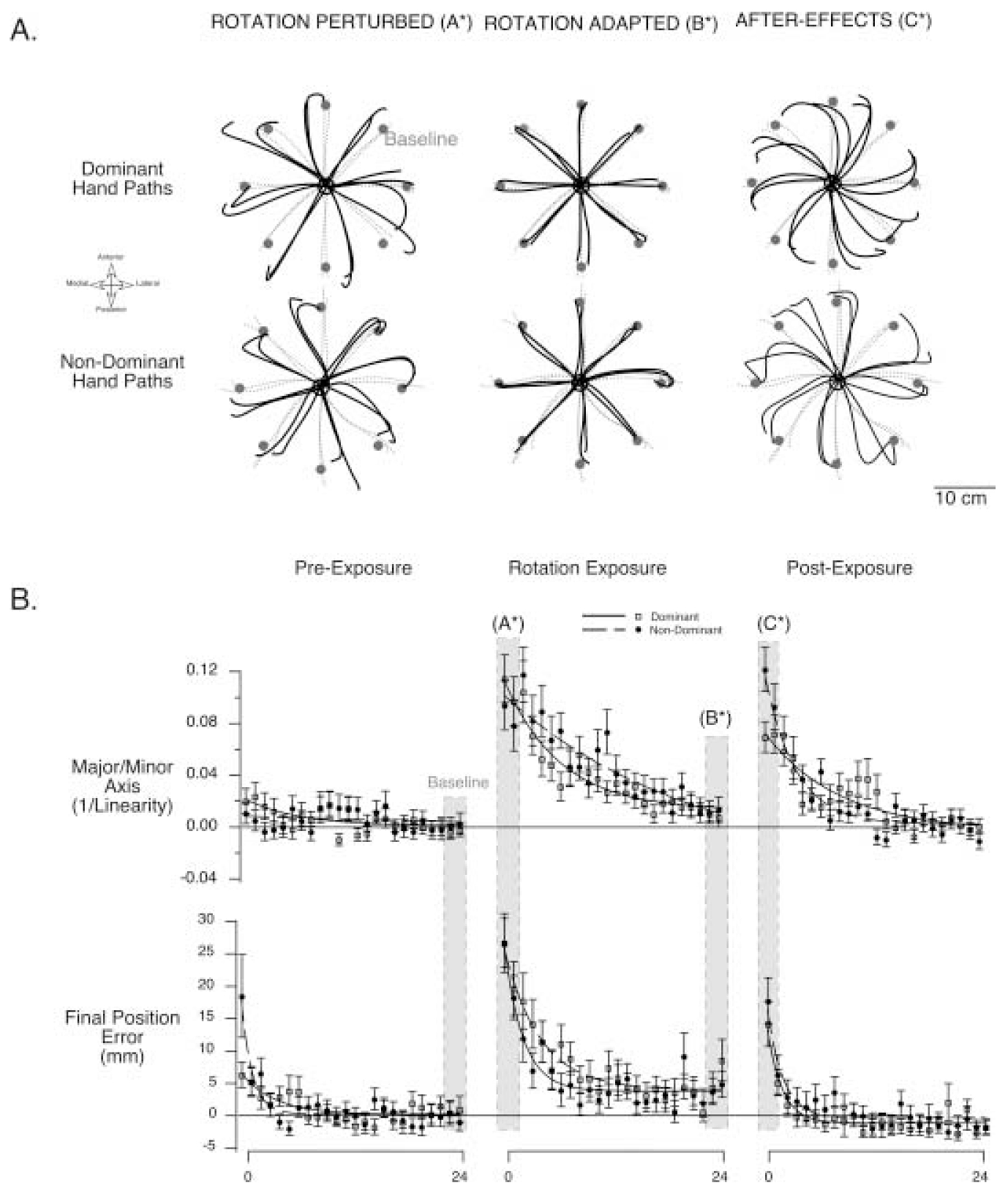
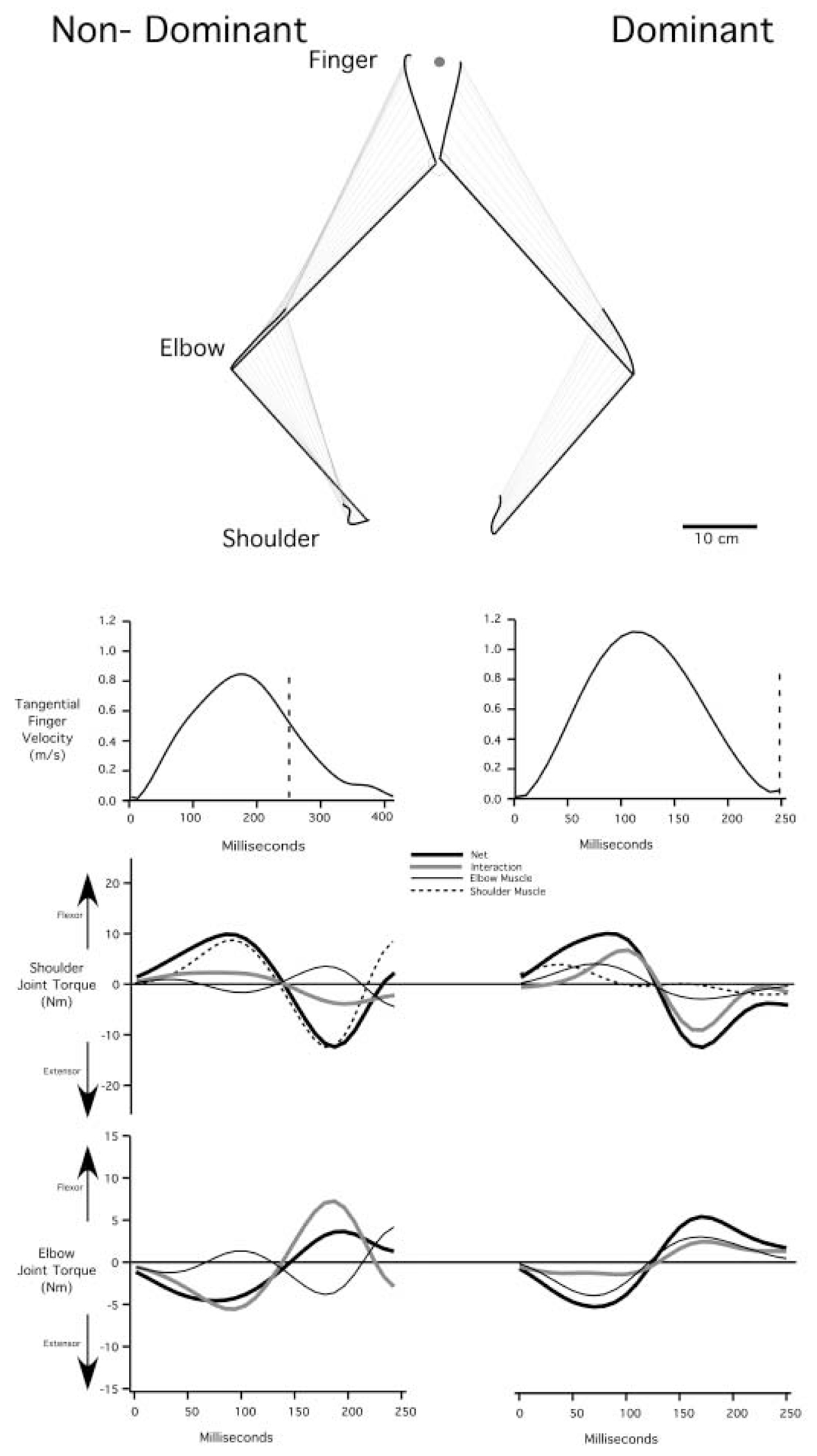
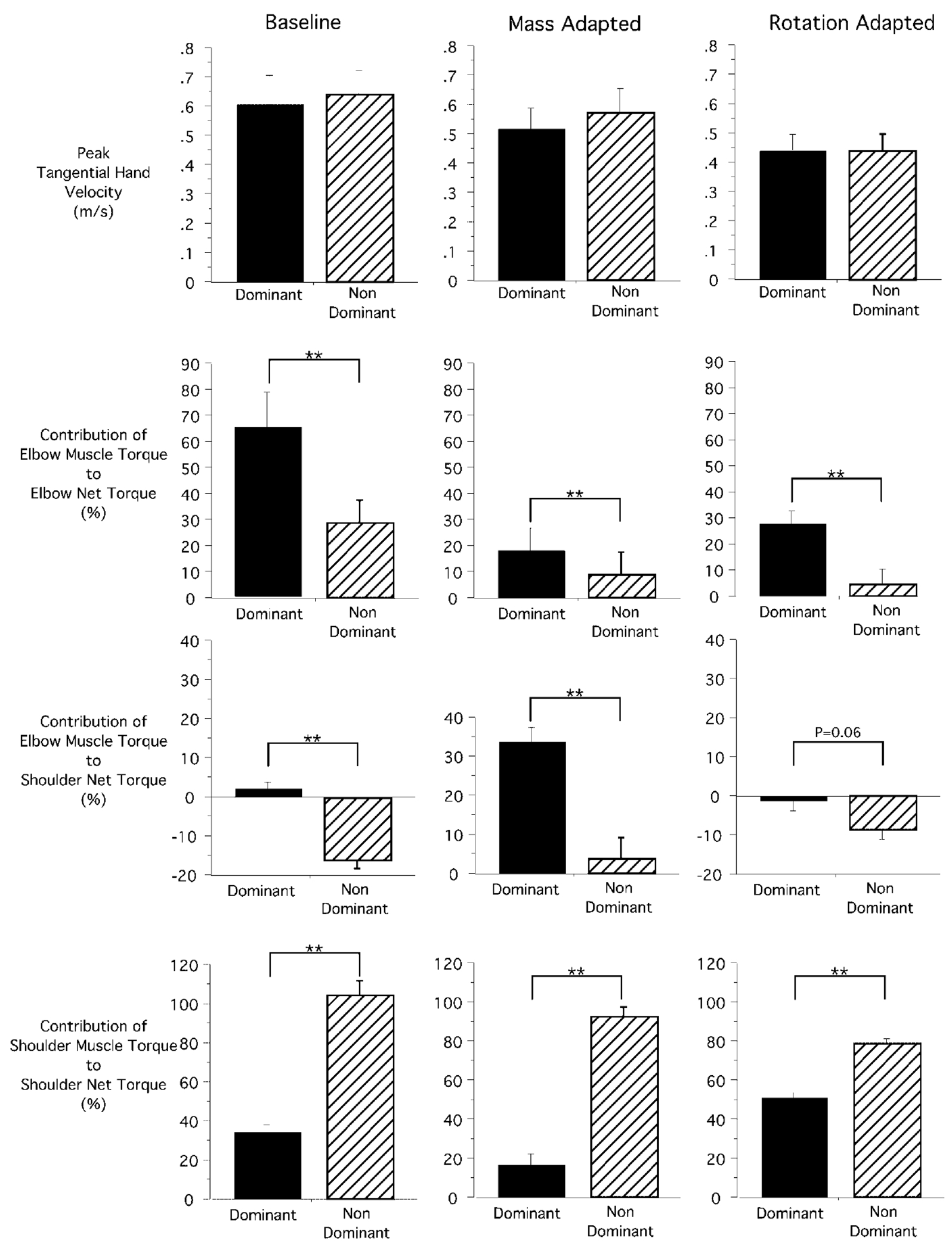
Handedness is a prominent behavioral phenomenon that emerges from asymmetrical neural organization of human motor systems. However, the aspects of motor performance that correspond to handedness remain largely undetermined. A recent study examining interlimb differences in coordination of reaching demonstrated dominant arm advantages in controlling limb segment inertial dynamics (Sainburg and Kalakanis 2000). Based on these findings, I now propose the dynamic-dominance hypothesis, which states that the essential factor that distinguishes dominant from nondominant arm performance is the facility governing the control of limb dynamics. The purpose of this study is to test two predictions of this hypothesis: 1) adaptation to novel intersegmental dynamics, requiring the development of new dynamic transforms, should be more effective for the dominant arm; 2) there should be no difference in adapting to visuomotor rotations performed with the dominant as compared with the nondominant arm. The latter prediction is based on the idea that visual information about target position is translated into an internal reference frame prior to transformation of the movement plan into dynamic properties, which reflect the forces required to produce movement. To test these predictions, dominant arm adaptation is compared to nondominant arm adaptation during exposure to novel inertial loads and to novel visuomotor rotations. The results indicate substantial interlimb differences in adaptation to novel inertial dynamics, but equivalent adaptation to novel visuomotor rotations. Inverse dynamic analysis revealed better coordination of dominant arm muscle torques across both shoulder and elbow joints, as compared with nondominant arm muscle torques. As a result, dominant arm movements were produced with a fraction of the mean squared muscle torque computed for nondominant arm movements made at similar speeds. These results support the dynamic-dominance hypothesis, indicating that interlimb asymmetries in control arise downstream to visuomotor transformations, when dynamic variables that correspond to the forces required for motion are specified.
Figures
References
-
- Abend W, Bizzi E, Morasso P (1982) Human arm trajectory formation. Brain 105:331–348 - PubMed
-
- Annett J, Annett M, Hudson PTW (1979) The control of movement in the preferred and non-preferred hands. Quart J Exp Psych 31:641–652 - PubMed
-
- Baily JS (1972) Adaptation to prisms: do proprioceptive changes mediate adapted behaviour with ballistic arm movements? Quart J Exp Psych 24:8–20 - PubMed
-
- Bentin S, Sahar A, Moscovitch M (1984) Intermanual information transfer in patients with lesions in the trunk of the corpus callosum. Neuropsych 22:601–611 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
