

Convergence of TOR-nitrogen and Snf1-glucose signaling pathways onto Gln3
- PMID: 11809814
- PMCID: PMC134645
- DOI: 10.1128/MCB.22.4.1246-1252.2002
Convergence of TOR-nitrogen and Snf1-glucose signaling pathways onto Gln3
Abstract
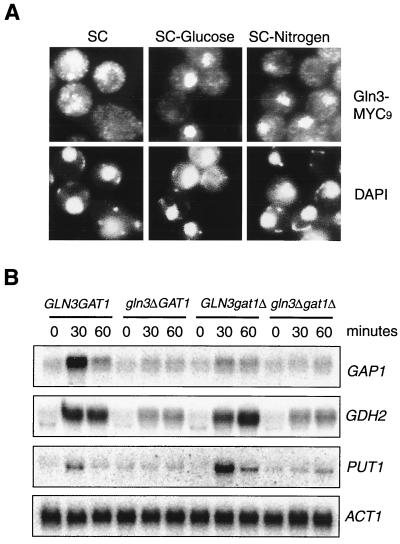
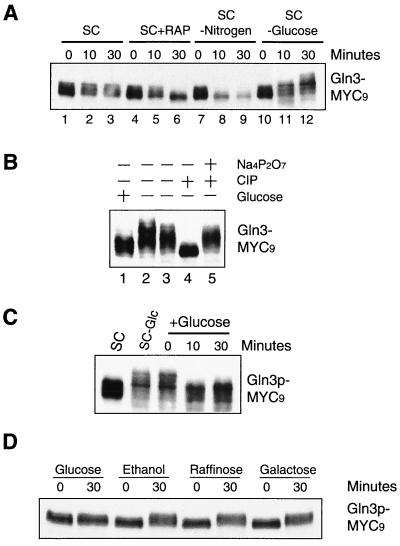
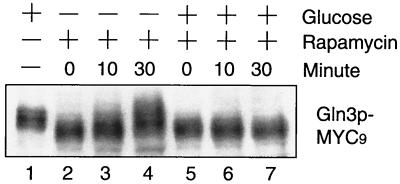
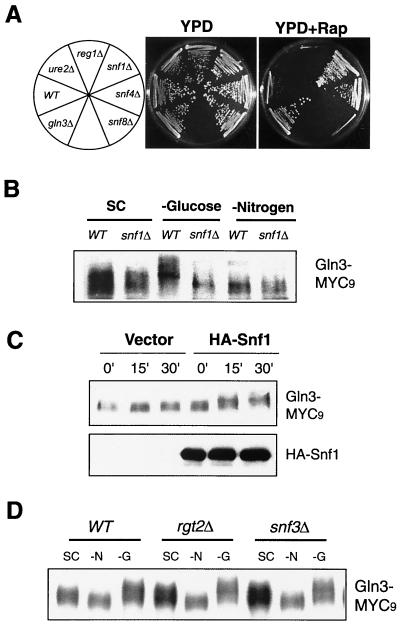
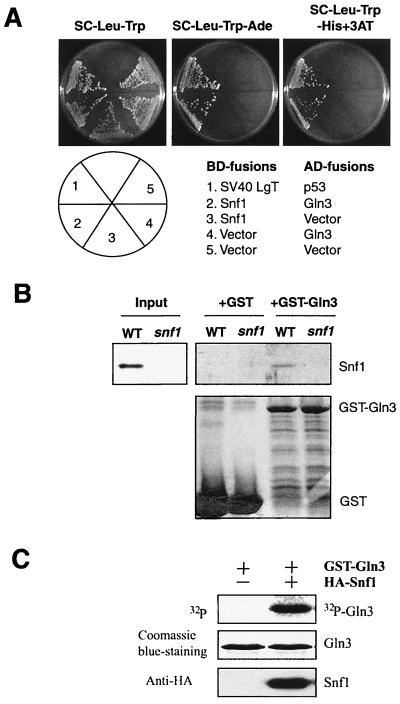
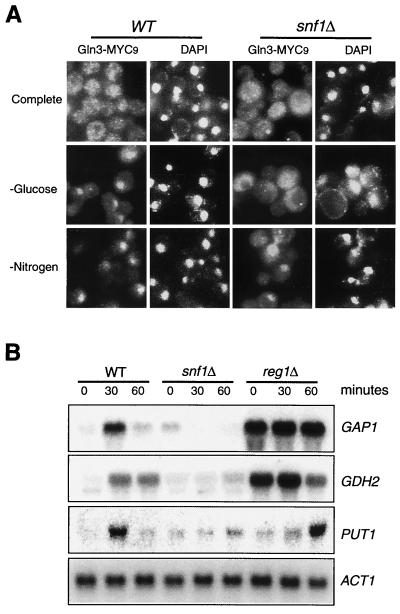
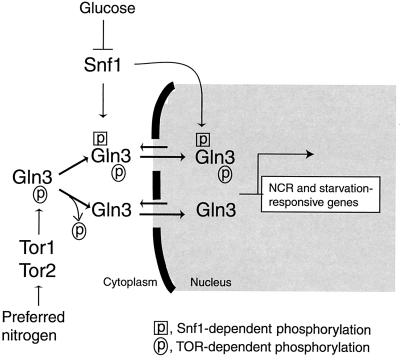
Carbon and nitrogen are two basic nutrient sources for cellular organisms. They supply precursors for energy metabolism and metabolic biosynthesis. In the yeast Saccharomyces cerevisiae, distinct sensing and signaling pathways have been described that regulate gene expression in response to the quality of carbon and nitrogen sources, respectively. Gln3 is a GATA-type transcription factor of nitrogen catabolite-repressible (NCR) genes. Previous observations indicate that the quality of nitrogen sources controls the phosphorylation and cytoplasmic retention of Gln3 via the target of rapamycin (TOR) protein. In this study, we show that glucose also regulates Gln3 phosphorylation and subcellular localization, which is mediated by Snf1, the yeast homolog of AMP-dependent protein kinase and a cytoplasmic glucose sensor. Our data show that glucose and nitrogen signaling pathways converge onto Gln3, which may be critical for both nutrient sensing and starvation responses.
Figures
References
-
- Alberts, B., D. Bray, J. Lewis, M. Raff, K. Roberts, and J. Watson. 1994. Molecular biology of the cell. Garland Publishing, Inc., New York, N.Y.
-
- Beck, T., and M. N. Hall. 1999. The TOR signaling pathway controls nuclear localization of nutrient-regulated transcriptional factors. Nature 402:689-692. - PubMed
-
- Bertram, P. G., J. Choi, J. Carvalho, W. D. Ai, C. B. Zeng, T. F. Chan, and X. F. S. Zheng. 2000. Tripartite regulation of Gln3p by TOR, Ure2p and phosphatases. J. Biol. Chem. 275:35727-35733. - PubMed
-
- Bertram, P. G., C. Zeng, J. Thorson, A. S. Shaw, and X. F. Zheng. 1998. The 14-3-3 proteins positively regulate rapamycin-sensitive signaling. Curr. Biol. 8:1259-1267. - PubMed
-
- Brachmann, C. B., A. Davies, G. J. Cost, E. Caputo, J. Li, P. Hieter, and J. D. Boeke. 1998. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast 14:115-132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases