

Action of prokaryotic enhancer over a distance does not require continued presence of promoter-bound sigma54 subunit
- PMID: 11809874
- PMCID: PMC100299
- DOI: 10.1093/nar/30.3.636
Action of prokaryotic enhancer over a distance does not require continued presence of promoter-bound sigma54 subunit
Abstract
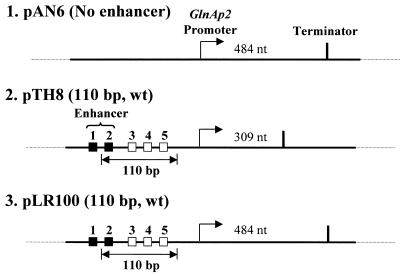
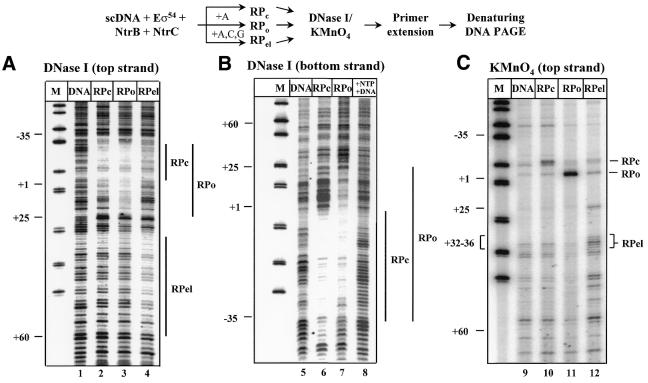
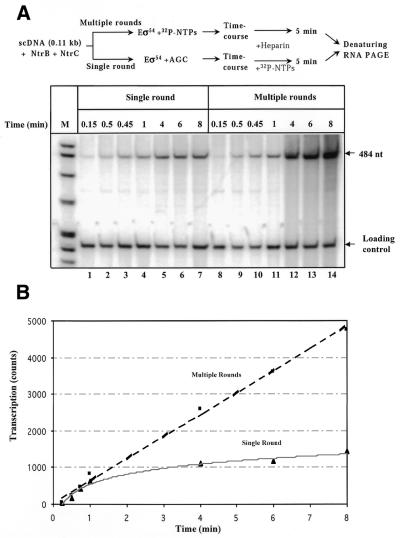
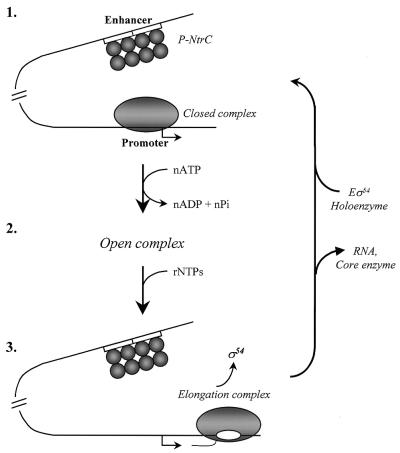
The mechanism by which an enhancer activates transcription over large distances has been investigated. Activation of the glnAp2 promoter by the NtrC-dependent enhancer in Escherichia coli was analyzed using a purified system supporting multiple-round transcription in vitro. Our results suggest that the enhancer-promoter interaction and the initiation complex must be formed de novo during every round of transcription. No protein remained bound to the promoter after RNA polymerase escaped into elongation. Furthermore, the rate of initiation during the first and subsequent rounds of transcription were very similar, suggesting that there was no functional 'memory' facilitating multiple rounds of transcription. These studies exclude the hypothesis that enhancer action during multiple-round transcription involves the memory of the initial activation event.
Figures

References
-
- Blackwood E.M. and Kadonaga,J.T. (1998) Going the distance: a current view of enhancer action. Science, 281, 61–63. - PubMed
-
- Magasanik B. (1993) The regulation of nitrogen utilization in enteric bacteria. J. Cell. Biochem., 51, 34–40. - PubMed
-
- Gralla J.D. (1996) Activation and repression of E. coli promoters. Curr. Opin. Genet. Dev., 6, 526–530. - PubMed
-
- Studholme D.J. and Buck,M. (2000) The biology of enhancer-dependent transcriptional regulation in bacteria: insights from genome sequences. FEMS Microbiol. Lett., 186, 1–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
