

doi: 10.1093/nar/29.24.5107.
An architectural role of the Escherichia coli chromatin protein FIS in organising DNA
Affiliations
- PMID: 11812843
- PMCID: PMC97572
- DOI: 10.1093/nar/29.24.5107
Item in Clipboard
An architectural role of the Escherichia coli chromatin protein FIS in organising DNA
Nucleic Acids Res.
.
Abstract
The Escherichia coli chromatin protein FIS modulates the topology of DNA in a growth phase-dependent manner. In this study we have investigated the global effect of FIS binding on DNA architecture in vitro. We show that in supercoiled DNA molecules FIS binds at multiple sites in a non-random fashion and increases DNA branching. This global DNA reshaping effect is independent of the helical phasing of FIS binding sites. We propose, in addition to the previously inferred stabilisation of tightly bent DNA microloops in the upstream regions of certain promoters, that FIS may perform the distinct architectural function of organising branched plectonemes in the E.coli nucleoid.
Figures
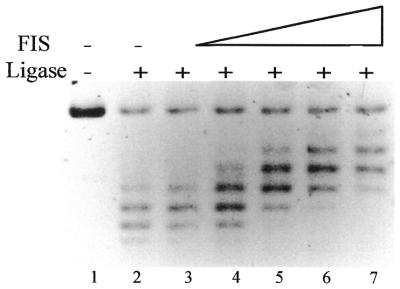
FIS introduces negative supercoils in DNA. Lane 1, open circular pBR322 DNA substrate; lane 2, only ligase was added. The FIS to DNA ratio in lanes 3–7 was ∼1 dimer per 800, 325, 160, 80 and 40 bp respectively. Under the conditions of electrophoresis used the more negatively supercoiled topoisomers migrate more slowly.
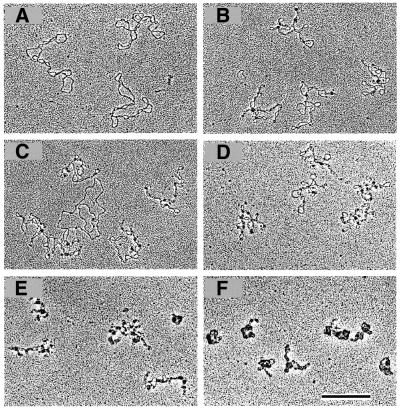
Binding of FIS leads to branching of supercoiled ptyrTlac DNA. EM images are shown. (A) Free supercoiled ptyrTlac DNA. FIS stabilises branches at a FIS to DNA ratio of ∼1 dimer per 325 bp DNA (B and C). Note that FIS, in agreement with a previous report (21), does not bind efficiently the relaxed (presumably nicked) DNA molecule in (C). The FIS to DNA ratio in (D), (E) and (F) was ∼1 dimer per 160, 80 and 40 bp, respectively. Note the ‘tightening’ of plasmid molecules with increasing FIS concentrations and the formation of compact structures at higher FIS to DNA ratios (scale bar represents 1 kb).
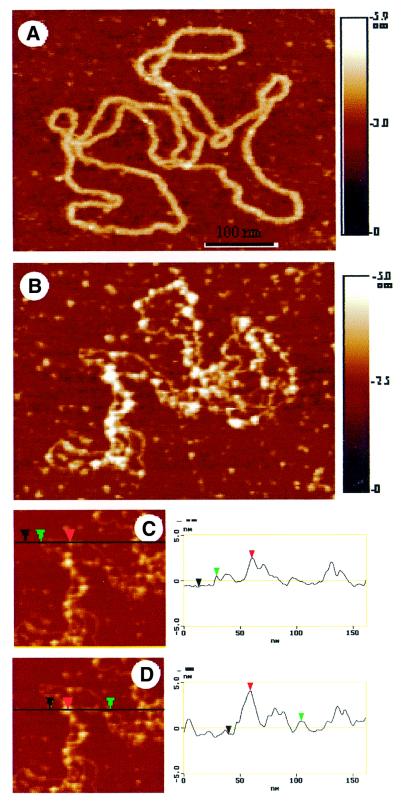
FIS forms clusters on binding supercoiled ptyrTlac DNA. AFM images are shown. (A) Free supercoiled ptyrTlac. (B) Complex formed at the FIS to ptyrTlac DNA ratio of ∼1 dimer per 80–100 bp DNA. (C and D) The profile plots are shown. The height of the DNA in these images is 1.3 ± 0.1 nm. The height of the protein is deduced from the height of a DNA–protein complex formed on a portion of the plasmid without crossover. The height of the protein is then 2.0 ± 0.1 nm. Considering this height of the protein and the height of the DNA, it is possible to analyse a crossover. This is shown in the profile plot (D). The height of the crossover (4.7 ± 0.2 nm) corresponds to the doubling of the DNA height and also the height of the protein. An apparent difference in the width of the DNA between (A) and (B) is due to a difference in the tip radii. Because AFM images are always a convolution between the tip shape and the real topography, the measured size of small features (in the range of the tip size) will vary from tip to tip.

FIS is associated with crossovers and branch points in pUC18 DNA. AFM images are shown (A). An overview of free pUC18 DNA molecules. The picture shown represents baseline corrected raw data, with no filtering. (B) A complex formed at the FIS to pUC18 DNA ratio of ∼1 dimer per 325 bp DNA. The image shown represents a three-dimensional rendering of baseline corrected raw data.
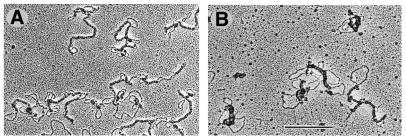
Electron micrographs of complexes of H-NS with ptyrTlac DNA. Elongated complexes (filaments) are formed presumably containing two DNA duplexes. Ratio of H-NS to DNA was ∼1 molecule per 10 bp (A) and ∼1 molecule per 3 bp (B). The scale bar represents 1 kb.
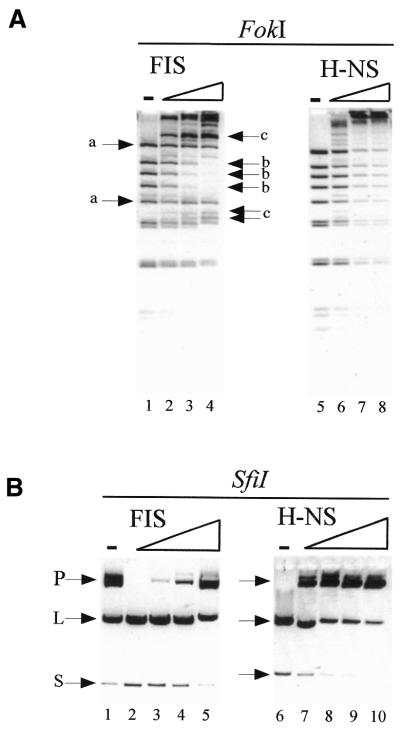
Probing of protein–DNA complexes with restricrtion endonucleases. (A) Probing with FokI. The FIS to DNA ratio in lanes 2–4 corresponded to 1 FIS dimer per 325, 100 and 40 bp, respectively. The H-NS to DNA ratio in lanes 6–8 corresponded to 1 molecule per 100, 30 and 10 bp, respectively. Electrophoresis was in 3% agarose gels. (A) Probing with SfiI. Low amounts of SfiI were used in the left panel (see lane 1) to reveal the stimulation of cleavage by FIS. The FIS to DNA ratio in lanes 2–5 corresponded to 1 FIS dimer per 325, 160, 80 and 40 bp, respectively. A 2-fold higher concentration of SfiI was used in the right panel (compare lanes 1 and 6) to reveal the inhibition of cleavage by H-NS. The H-NS to DNA ratio in lanes 7–10 corresponded to 1 molecule per 300, 100, 30 and 10 bp, respectively. The substrate plasmid DNA (P), the large (L) and 900 bp small (S) cleavage products are indicated. Electrophoresis was in 1% agarose gels.
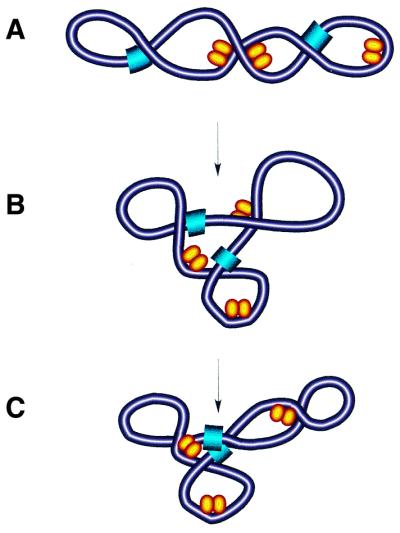
Model of DNA reshaping by FIS. (A) Binding and bending of isolated DNA regions in plectonemically supercoiled DNA by single FIS dimers (yellow ellipsoids) facilitates the branching of DNA with resultant multiplication of apical loops. (B) Note that the FIS-dependent branching of DNA locates the two remote sites (blue) in close spatial proximity (C).
References
-
- Sinden R.R., Carlson,J.O. and Pettijohn,D.E. (1980) Torsional tension in the DNA double helix measured with trimethylpsoralen in living E. coli cells: analogous measurements in insect and human cells. Cell, 21, 773–783. - PubMed
-
- Boles T.C., White,J.H. and Cozzarelli,N.R. (1990) Structure of plectonemically supercoiled DNA. J. Mol. Biol., 213, 931–951. - PubMed
-
- Worcel A. and Burgi,E. (1972) On the structure of the folded chromosome of Escherichia coli.J. Mol. Biol., 71, 127–147. - PubMed
-
- Staczek P. and Higgins,N.P. (1998) Gyrase and topo IV modulate chromosome domain size in vivo. Mol. Microbiol., 29, 1435–1448. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
