

doi: 10.1093/emboj/21.3.355.
Identification of a ubiquitin-protein ligase subunit within the CCR4-NOT transcription repressor complex
Affiliations
- PMID: 11823428
- PMCID: PMC125831
- DOI: 10.1093/emboj/21.3.355
Item in Clipboard
Identification of a ubiquitin-protein ligase subunit within the CCR4-NOT transcription repressor complex
EMBO J.
.
Abstract
The RING finger protein CNOT4 is a component of the CCR4-NOT complex. This complex is implicated in repression of RNA polymerase II transcription. Here we demonstrate that CNOT4 functions as a ubiquitin-protein ligase (E3). We show that the unique C4C4 RING domain of CNOT4 interacts with a subset of ubiquitin-conjugating enzymes (E2s). Using NMR spectroscopy, we detail the interaction of CNOT4 with UbcH5B and characterize RING residues that are critical for this interaction. CNOT4 acts as a potent E3 ligase in vitro. Mutations that destabilize the E2-E3 interface abolish this activity. Based on these results, we present a model of how E3 ligase function within the CCR4-NOT complex relates to transcriptional regulation.
Figures
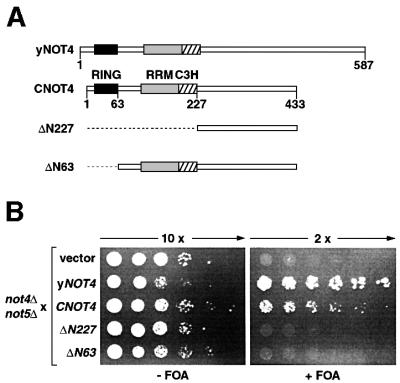
Fig. 1. Complementation analysis of yeast strain YOU637 (not4Δ not5Δ). (A) Schematic representation of yNOT4, CNOT4 and CNOT4 derivatives. The RING, RRM and C3H zinc finger domains are shown as black, gray and hatched boxes, respectively. Truncations of ΔN227 and ΔN63 are indicated by stippled lines. (B) YOU637 cells were transformed with empty vector, yNOT4 and CNOT4 full-length and partial cDNAs as indicated. Ten- and 2-fold serial dilutions of transformants were spotted on selective medium in either the absence (–) or presence (+) of 0.1% FOA, and colonies were grown for 4 days at 30°C.
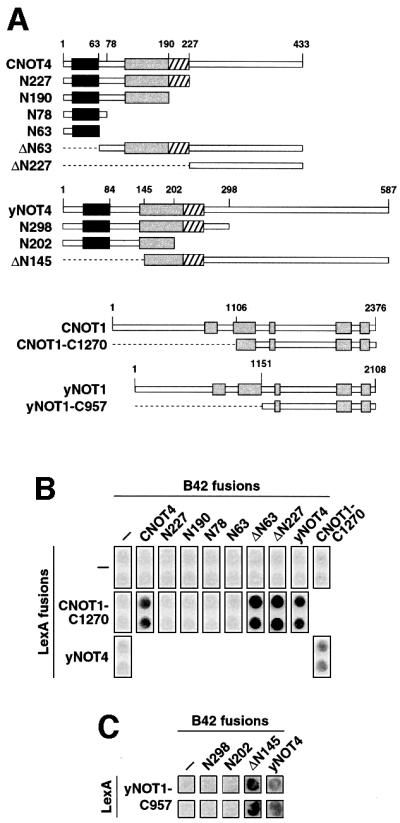
Fig. 2. CNOT4 interacts with CNOT1. (A) Survey of CNOT4, yNOT4, CNOT1 and yNOT1 derivatives used in two-hybrid assays. Conserved motifs within NOT1 proteins are indicated by gray shading. (B) Two-hybrid interactions of CNOT4 with CNOT1. EGY48 yeast cells containing a LexA-responsive lacZ reporter gene were transformed with the indicated LexA and B42 fusion plasmids. Empty vector controls (–) were included. Transformants were assayed on X-gal plates for blue staining. (C) Two-hybrid interactions of yNOT4 with yNOT1. Analysis was performed as in (B).
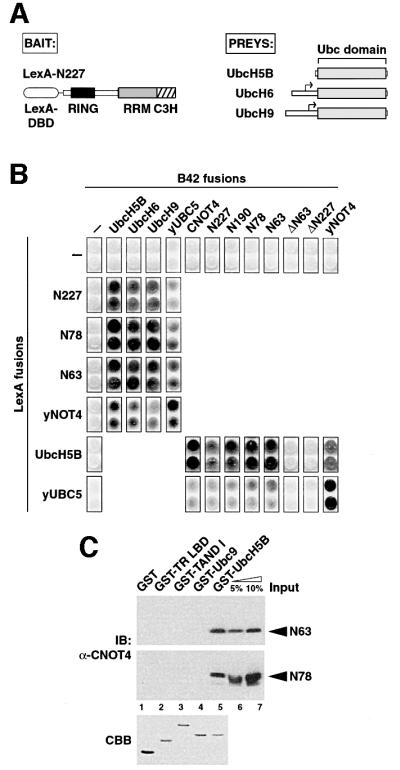
Fig. 3. Specific interactions of CNOT4 with ubiquitin-conjugating E2 enzymes. (A) Schematic illustrations of two-hybrid ‘bait’ LexA-N227 (left) and of E2s isolated as ‘preys’ (right). Small arrows denote start codons of the partial UbcH6 and UbcH9 library clones. (B) (C)NOT4 interactions with human and yeast Ubc proteins in the two-hybrid system. (C) GST pull-down analysis of His6-N63 and His6-N78 RING finger proteins using different GST fusion proteins: lane 1, GST alone; lane 2, GST fusions with the ligand-binding domain of chicken thyroid receptor α (GST-TR LBD); lane 3, Drosphila TAFII230 N-terminal domain (GST-TAND I); lane 4, SUMO-1-conjugating enzyme Ubc9 (GST-Ubc9); and lane 5, GST–UbcH5B. The upper and middle panels show immunoblots (IB) using anti-CNOT4 antibody 19A12. Five and ten percent of recombinant N63 and N78 used in the pull-down reactions were loaded as input controls (lanes 6 and 7). The lower panel shows a Coomassie Blue (CBB) stain of an SDS–gel with 2 µg of each GST protein loaded.
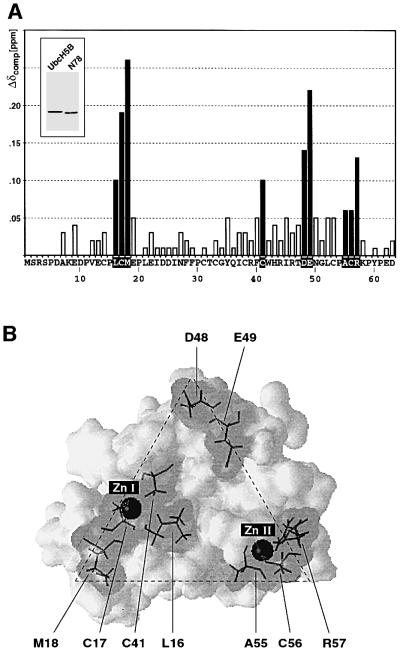
Fig. 4. Titration of UbcH5B with CNOT4 RING. (A) Compound chemical shifts (Δδcomp) of 15N-labeled His6-N78 upon titration of UbcH5B. Residues showing chemical shift changes >0.05 p.p.m. are shown as black bars. The inset shows a Coomassie Blue-stained gel of UbcH5B (lane 1) and His6-N78 (lane 2). Only shifts for residues 7–63 could be detected in 15N-HSQC spectra. (B) Three-dimensional surface representation of CNOT4 RING structure. Residues showing shifts >0.05 p.p.m. are indicated by gray shading. Stippled lines outline the triangular shape of the E2 interface.
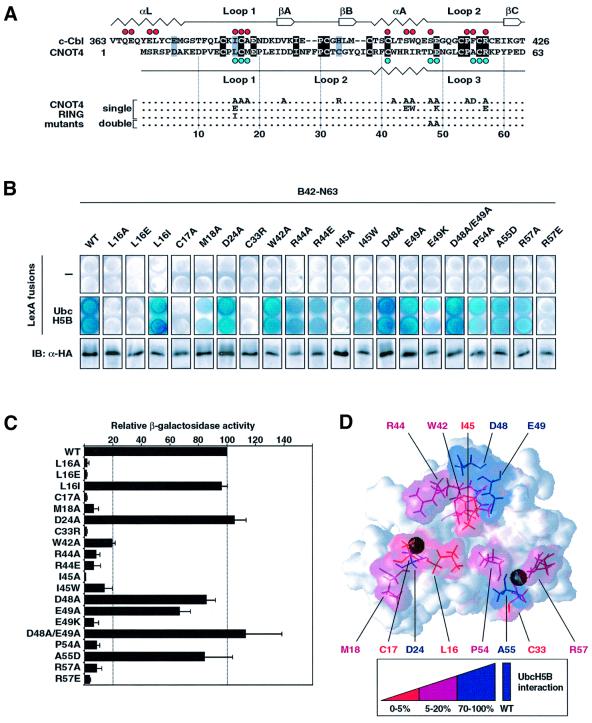
Fig. 5. Effect of CNOT4 RING mutations on UbcH5B interaction. (A) Survey of CNOT4 RING mutations. An alignment of CNOT4 and the c-Cbl RING sequences is shown on the top, with secondary structure elements displayed below and above the sequences, respectively. Red dots indicate contacts of the c-Cbl–UbcH7 interface (Zheng et al., 2000), those in blue indicate CNOT4 chemical shifts >0.05 p.p.m. (B) Yeast strains expressing empty (–) vector (upper panels) or LexA-UbcH5B (middle panels) were transformed with HA-tagged wild type (WT) and mutant B42-N63 as indicated. Equal expression levels of all B42 fusion proteins were confirmed by anti-HA immunoblot analysis (lower panels). Individual clones were assayed on X-gal plates (B), and by quantitative determination of the means and SEM of relative β-galactosidase activities in yeast cell extracts, with B42-N63 (WT) set to 100% (C). (D) Three-dimensional map illustrating the effects of mutations on UbcH5B binding by color coding: abolishment, red; strongly affected, magenta; not affected, blue.
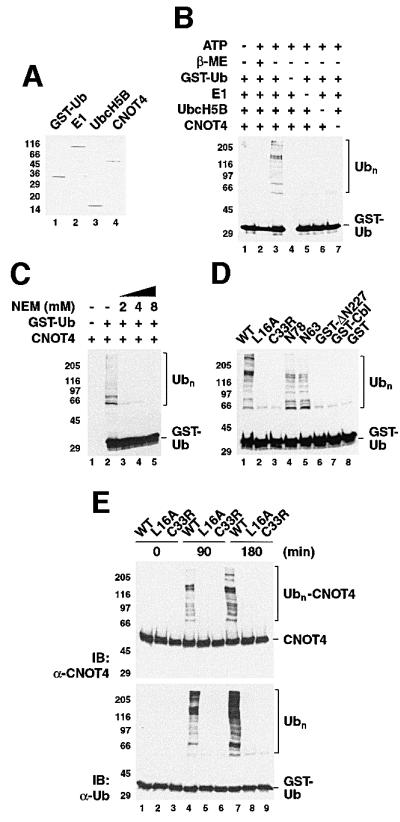
Fig. 6. E3 ligase activity of CNOT4 in vitro. (A) Coomassie Blue-stained gel showing components of the in vitro ubiquitylation system: lane 1, GST–ubiquitin; lane 2, ubiquitin-activating enzyme E1; lane 3, UbcH5B; and lane 4, His6-CNOT4. (B) Ubiquitylation reactions were performed for 90 min in the absence (–) or presence (+) of 2 mM ATP with the indicated components. In lane 2, β-mercaptoethanol (β-ME) was included to 2 M. The western blot used anti-ubiquitin antibody. Polyubiquitin chains (Ubn) are bracketed. (C) Effect of the thiol-specific reagent N-ethylmaleimide (NEM) on Ubn formation. (D) Ubiquitylation reactions used wild-type (WT) and mutant His6-CNOT4, N78 and N63 proteins, or GST fusion proteins as indicated. GST–Cbl denotes a fusion of c-Cbl residues 47–447. The reactions were performed for 90 min (left panel), and the reaction products probed with anti-ubiquitin antibody. (E) Ubiquitylation reactions using wild-type (WT) and mutant (L16A, C33R) His6-CNOT4 were first analyzed with anti-CNOT4 (upper panel) and reprobed with anti-ubiquitin antibody (lower panel).
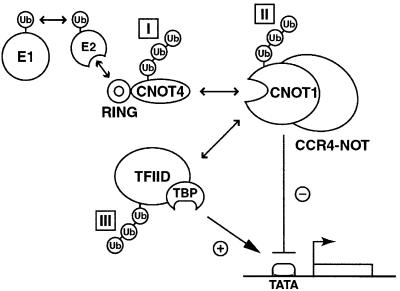
Fig. 7. Model illustrating potential targets for RING-dependent CNOT4 E3 ligase activity: I, CNOT4 itself; II, CNOT1 or other CCR4–NOT subunits; and III, TFIID. Arrows indicate physical and functional connections. See text for details.
References
-
- Albright S.R. and Tjian,R. (2000) TAFs revisited: more data reveal twists and confirm old ideas. Gene, 242, 1–13. - PubMed
-
- Barlow P.N., Luisi,B., Milner,A., Elliott,M. and Everett,R. (1994) Structure of the C3HC4 domain by 1H-nuclear magnetic resonance spectroscopy. A new structural class of zinc-finger. J. Mol. Biol., 237, 201–211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
