

Activation of the JNK pathway during dorsal closure in Drosophila requires the mixed lineage kinase, slipper
- PMID: 11825878
- PMCID: PMC155330
- DOI: 10.1101/gad.953002
Activation of the JNK pathway during dorsal closure in Drosophila requires the mixed lineage kinase, slipper
Abstract
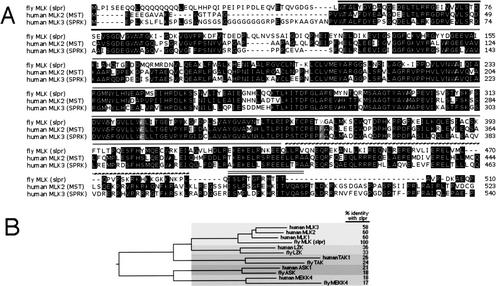
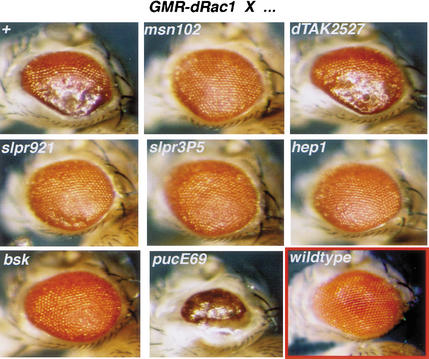
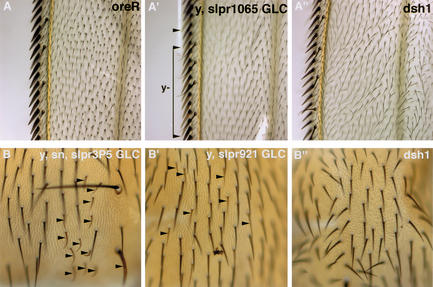
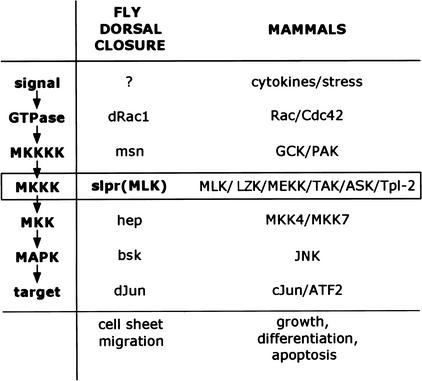
The Jun kinase (JNK) pathway has been characterized for its role in stimulating AP-1 activity and for modulating the balance between cell growth and death during development, inflammation, and cancer. Six families of mammalian kinases acting at the level of JNKKK have emerged as upstream regulators of JNK activity (MLK, LZK, TAK, ASK, MEKK, and TPL); however, the specificity underlying which kinase is utilized for transducing a distinct signal is poorly understood. In Drosophila, JNK signaling plays a central role in dorsal closure, controlling cell fate and cell sheet morphogenesis during embryogenesis. Notably, in the fly genome, there are single homologs of each of the mammalian JNKKK families. Here, we identify mutations in one of those, a mixed lineage kinase, named slipper (slpr), and show that it is required for JNK activation during dorsal closure. Furthermore, our results show that other putative JNKKKs cannot compensate for the loss of slpr function and, thus, may regulate other JNK or MAPK-dependent processes.
Figures



Similar articles
-
Drosophila mixed lineage kinase/slipper, a missing biochemical link in Drosophila JNK signaling.Biochim Biophys Acta. 2003 Apr 7;1640(1):77-84. doi: 10.1016/s0167-4889(03)00022-3. Biochim Biophys Acta. 2003. PMID: 12676357
-
The role of the Drosophila TAK homologue dTAK during development.Mech Dev. 2001 Apr;102(1-2):67-79. doi: 10.1016/s0925-4773(01)00285-4. Mech Dev. 2001. PMID: 11287182
-
Genetic analysis of slipper/mixed lineage kinase reveals requirements in multiple Jun-N-terminal kinase-dependent morphogenetic events during Drosophila development.Genetics. 2006 Oct;174(2):719-33. doi: 10.1534/genetics.106.056564. Epub 2006 Aug 3. Genetics. 2006. PMID: 16888342 Free PMC article.
-
Mixed-lineage kinase control of JNK and p38 MAPK pathways.Nat Rev Mol Cell Biol. 2002 Sep;3(9):663-72. doi: 10.1038/nrm906. Nat Rev Mol Cell Biol. 2002. PMID: 12209126 Review.
-
Regulating cell morphogenesis: the Drosophila Jun N-terminal kinase pathway.Genesis. 2013 Mar;51(3):147-62. doi: 10.1002/dvg.22354. Epub 2012 Nov 20. Genesis. 2013. PMID: 23109363 Review.
Cited by
-
Characterization of TNF-induced cell death in Drosophila reveals caspase- and JNK-dependent necrosis and its role in tumor suppression.Cell Death Dis. 2019 Aug 14;10(8):613. doi: 10.1038/s41419-019-1862-0. Cell Death Dis. 2019. PMID: 31409797 Free PMC article.
-
The HIV-1 Vpu protein induces apoptosis in Drosophila via activation of JNK signaling.PLoS One. 2012;7(3):e34310. doi: 10.1371/journal.pone.0034310. Epub 2012 Mar 29. PLoS One. 2012. PMID: 22479597 Free PMC article.
-
Role of MLK3 in the regulation of mitogen-activated protein kinase signaling cascades.Mol Cell Biol. 2005 May;25(9):3670-81. doi: 10.1128/MCB.25.9.3670-3681.2005. Mol Cell Biol. 2005. PMID: 15831472 Free PMC article.
-
The PP2C Alphabet is a negative regulator of stress-activated protein kinase signaling in Drosophila.Genetics. 2009 Feb;181(2):567-79. doi: 10.1534/genetics.108.096461. Epub 2008 Dec 8. Genetics. 2009. PMID: 19064708 Free PMC article.
-
Rho1 regulates apoptosis via activation of the JNK signaling pathway at the plasma membrane.J Cell Biol. 2010 Apr 19;189(2):311-23. doi: 10.1083/jcb.200912010. J Cell Biol. 2010. PMID: 20404112 Free PMC article.
References
-
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, Amanatides PG, Scherer SE, Li PW, Hoskins RA, Galle RF, et al. The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. - PubMed
-
- Bock BC, Vacratsis PO, Qamirani E, Gallo KA. Cdc42-induced activation of the mixed-lineage kinase SPRK in vivo. Requirement of the Cdc42/Rac interactive binding motif and changes in phosphorylation. J Biol Chem. 2000;275:14231–14241. - PubMed
-
- Boutros M, Paricio N, Strutt DI, Mlodzik M. Dishevelled activates JNK and discriminates between JNK pathways in planar polarity and wingless signaling. Cell. 1998;94:109–118. - PubMed
-
- Burbelo PD, Drechsel D, Hall A. A conserved binding motif defines numerous candidate target proteins for both Cdc42 and Rac GTPases. J Biol Chem. 1995;270:29071–29074. - PubMed
-
- Campos-Ortega JA, Hartenstein V. The embryonic development of Drosophila melanogaster. Berlin/New York: Springer; 1997.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous