

Dipeptidyl peptidase I activates neutrophil-derived serine proteases and regulates the development of acute experimental arthritis
- PMID: 11827996
- PMCID: PMC150852
- DOI: 10.1172/JCI13462
Dipeptidyl peptidase I activates neutrophil-derived serine proteases and regulates the development of acute experimental arthritis
Abstract
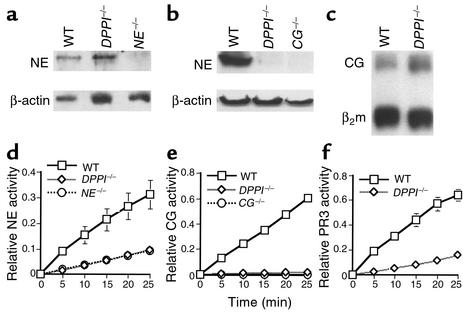
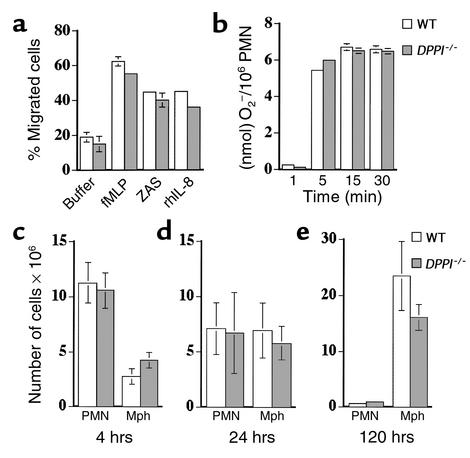
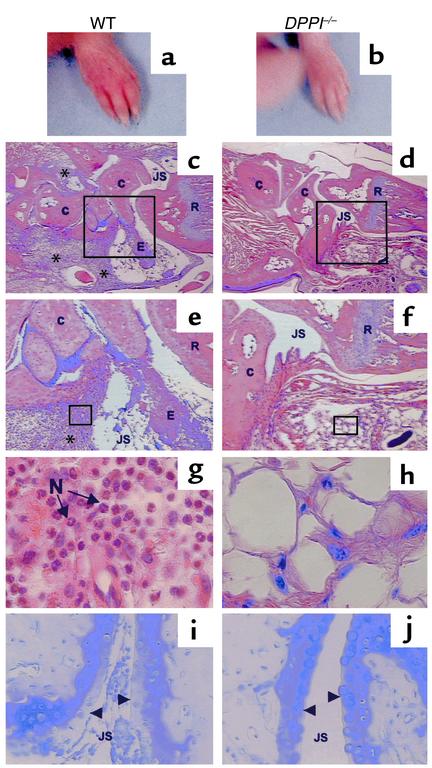
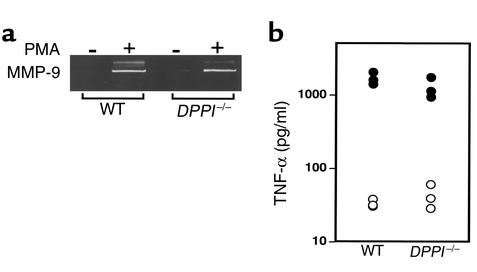
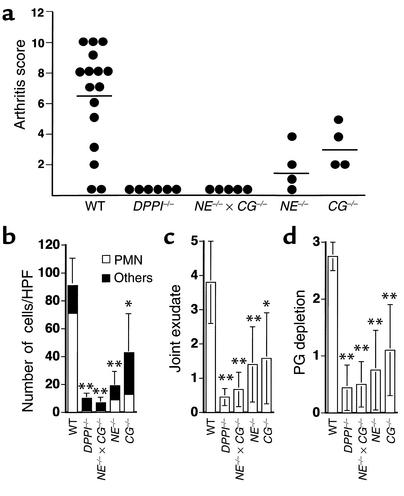
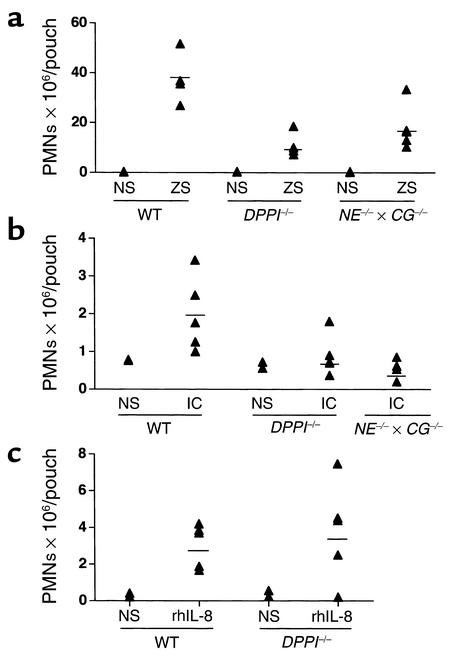
Leukocyte recruitment in inflammation is critical for host defense, but excessive accumulation of inflammatory cells can lead to tissue damage. Neutrophil-derived serine proteases (cathepsin G [CG], neutrophil elastase [NE], and proteinase 3 [PR3]) are expressed specifically in mature neutrophils and are thought to play an important role in inflammation. To investigate the role of these proteases in inflammation, we generated a mouse deficient in dipeptidyl peptidase I (DPPI) and established that DPPI is required for the full activation of CG, NE, and PR3. Although DPPI(-/-) mice have normal in vitro neutrophil chemotaxis and in vivo neutrophil accumulation during sterile peritonitis, they are protected against acute arthritis induced by passive transfer of monoclonal antibodies against type II collagen. Specifically, there is no accumulation of neutrophils in the joints of DPPI(-/-) mice. This protective effect correlates with the inactivation of neutrophil-derived serine proteases, since NE(-/-) x CG(-/-) mice are equally resistant to arthritis induction by anti-collagen antibodies. In addition, protease-deficient mice have decreased response to zymosan- and immune complex-mediated inflammation in the subcutaneous air pouch. This defect is accompanied by a decrease in local production of TNF-alpha and IL-1 beta. These results implicate DPPI and polymorphonuclear neutrophil-derived serine proteases in the regulation of cytokine production at sites of inflammation.
Figures
References
-
- Lehrer RI, Ganz T. Antimicrobial polypeptides of human neutrophils. Blood. 1990;76:2169–2181. - PubMed
-
- Chertov O, Yang D, Howard OM, Oppenheim JJ. Leukocyte granule proteins mobilize innate host defenses and adaptive immune responses. Immunol Rev. 2000;177:68–78. - PubMed
-
- Weiss SJ. Tissue destruction by neutrophils. N Engl J Med. 1989;320:365–376. - PubMed
-
- Lehr H, Arfors K. Mechanisms of tissue damage by leukocytes. Curr Opin Hematol. 1994;1:92–99. - PubMed
-
- Caughey GH. Serine proteinases of mast cell and leukocyte granules. A league of their own. Am J Respir Crit Care Med. 1994;150:S138–S142. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
