

Regulation of the tyrosine kinase Itk by the peptidyl-prolyl isomerase cyclophilin A
- PMID: 11830645
- PMCID: PMC122291
- DOI: 10.1073/pnas.042529199
Regulation of the tyrosine kinase Itk by the peptidyl-prolyl isomerase cyclophilin A
Abstract
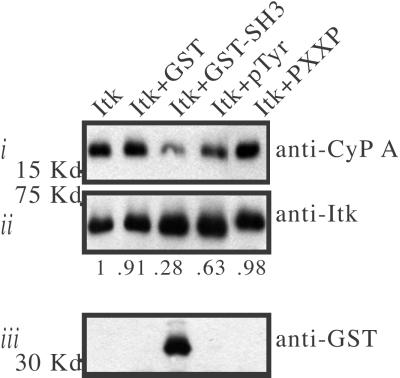
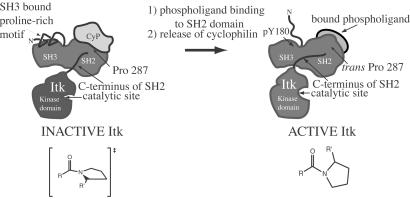
Interleukin-2 tyrosine kinase (Itk) is a nonreceptor protein tyrosine kinase of the Tec family that participates in the intracellular signaling events leading to T cell activation. Tec family members contain the conserved SH3, SH2, and catalytic domains common to many kinase families, but they are distinguished by unique sequences outside of this region. The mechanism by which Itk and related Tec kinases are regulated is not well understood. Our studies indicate that Itk catalytic activity is inhibited by the peptidyl prolyl isomerase activity of cyclophilin A (CypA). NMR structural studies combined with mutational analysis show that a proline-dependent conformational switch within the Itk SH2 domain regulates substrate recognition and mediates regulatory interactions with the active site of CypA. CypA and Itk form a stable complex in Jurkat T cells that is disrupted by treatment with cyclosporin A. Moreover, the phosphorylation levels of Itk and a downstream substrate of Itk, PLCgamma1, are increased in Jurkat T cells that have been treated with cyclosporin A. These findings support a novel mode of tyrosine kinase regulation for a Tec family member and provide a molecular basis for understanding a cellular function of the ubiquitous peptidyl prolyl isomerase, CypA.
Figures
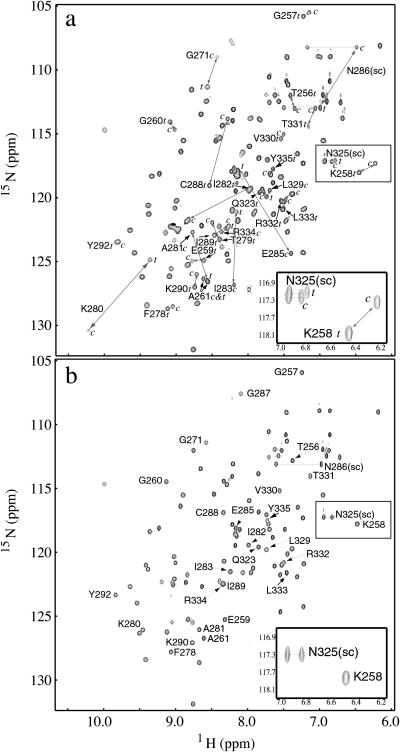
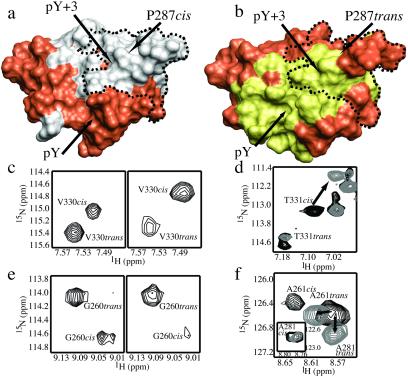
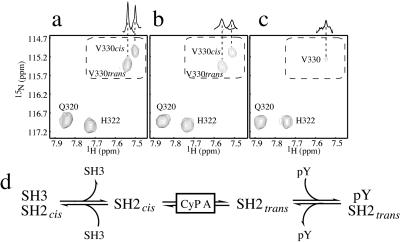
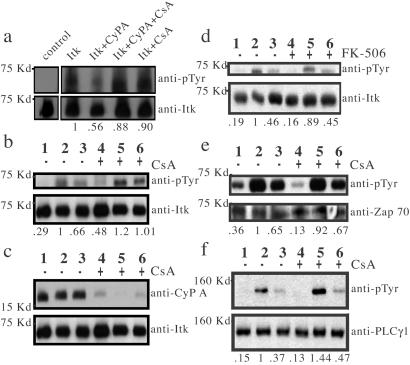
References
-
- Hubbard S R, Till J H. Annu Rev Biochem. 2000;69:373–398. - PubMed
-
- Xu W, Harrison S C, Eck M J. Nature (London) 1997;385:595–602. - PubMed
-
- Sicheri F, Moarefi I, Kuriyan J. Nature (London) 1997;385:602–609. - PubMed
-
- Willams J C, Wierenga R K, Saraste M. Trends Biochem Sci. 1998;23:179–184. - PubMed
-
- Yang W-C, Collette Y, Nunès J A, Olive D. Immunity. 2000;12:373–382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
