

Calcium signaling: a tale for all seasons
- PMID: 11830654
- PMCID: PMC122154
- DOI: 10.1073/pnas.032427999
Calcium signaling: a tale for all seasons
Abstract
An experiment performed in London nearly 120 years ago, which by today's standards would be considered unacceptably sloppy, marked the beginning of the calcium (Ca(2+)) signaling saga. Sidney Ringer [Ringer, S. (1883) J. Physiol. 4, 29-43] was studying the contraction of isolated rat hearts. In earlier experiments, Ringer had suspended them in a saline medium for which he admitted to having used London tap water, which is hard: The hearts contracted beautifully. When he proceeded to replace the tap water with distilled water, he made a startling finding: The beating of the hearts became progressively weaker, and stopped altogether after about 20 min. To maintain contraction, he found it necessary to add Ca(2+) salts to the suspension medium. Thus, Ringer had serendipitously discovered that Ca(2+), hitherto exclusively considered as a structural element, was active in a tissue that has nothing to do with bone or teeth, and performed there a completely novel function: It carried the signal that initiated heart contraction. It was a landmark observation, which should have immediately aroused wide interest. Unexpectedly, however, for decades it attracted no particular attention. Occasionally, farsighted pioneers argued forcefully for a messenger role of Ca(2+), offering compelling experimental evidence. Among them, one could quote L. V. Heilbrunn [Heilbrunn, L. V. (1940) Physiol. Zool. 13, 88-94], who contracted frog muscle fibers by applying Ca(2+) salts to their cut ends, but not to their surfaces. Heilbrunn correctly concluded that Ca(2+) had diffused from the cut ends to the internal contractile elements to elicit their contraction. One could also quote K. Bailey [Bailey, K. (1942) Biochem. J. 36, 121-139], who showed that the ATPase activity of myosin was strongly activated by Ca(2+) (but not by Mg(2+)), and concluded that the liberation of Ca(2+) in the neighborhood of the myosin controlled muscle contraction. Clearly, enough evidence was there, but only a handful of people had the vision to see it and to foresee its far-reaching implications. Perhaps no better example of clairvoyance can be offered than the quip by O. Loewy in 1959: "Ja Kalzium, das ist alles!"
Figures
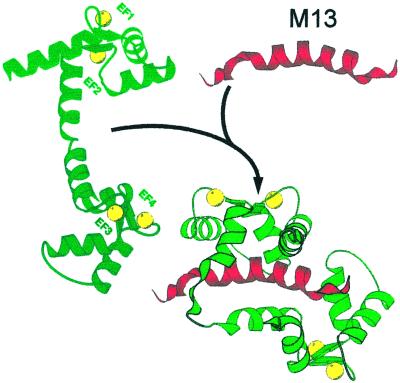
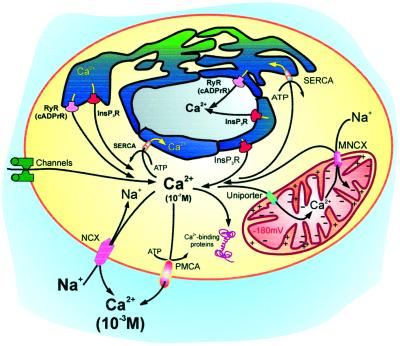
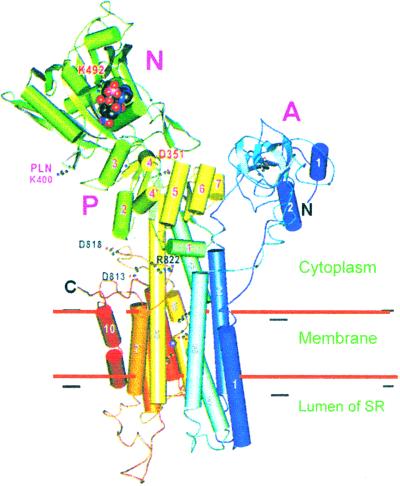
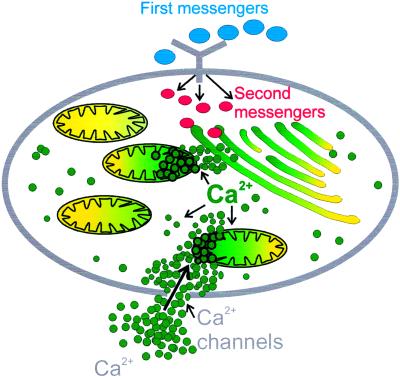
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
