

Ubiquitination of both adeno-associated virus type 2 and 5 capsid proteins affects the transduction efficiency of recombinant vectors
- PMID: 11836382
- PMCID: PMC135943
- DOI: 10.1128/jvi.76.5.2043-2053.2002
Ubiquitination of both adeno-associated virus type 2 and 5 capsid proteins affects the transduction efficiency of recombinant vectors
Abstract
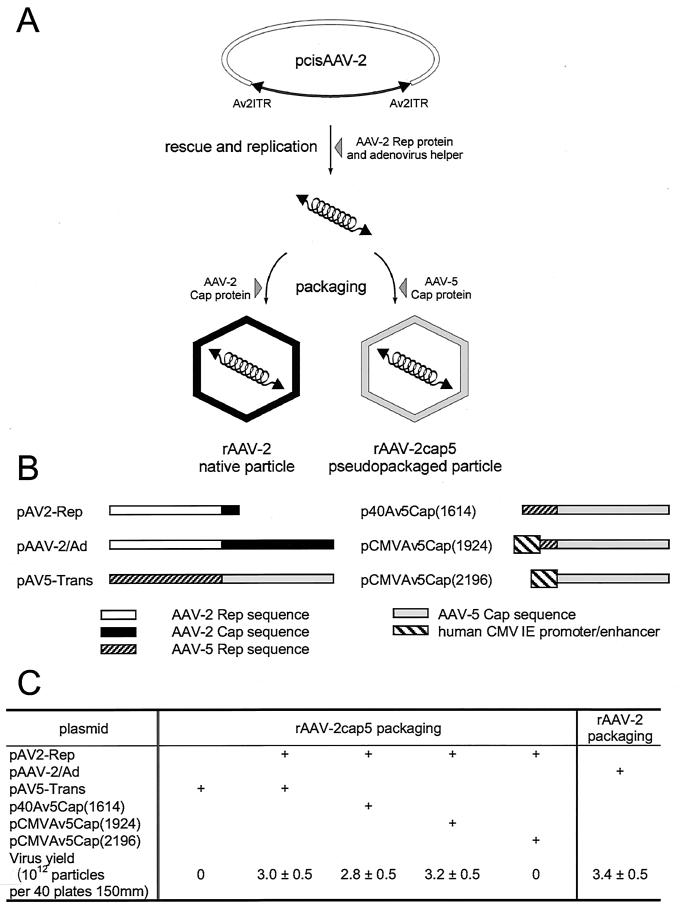
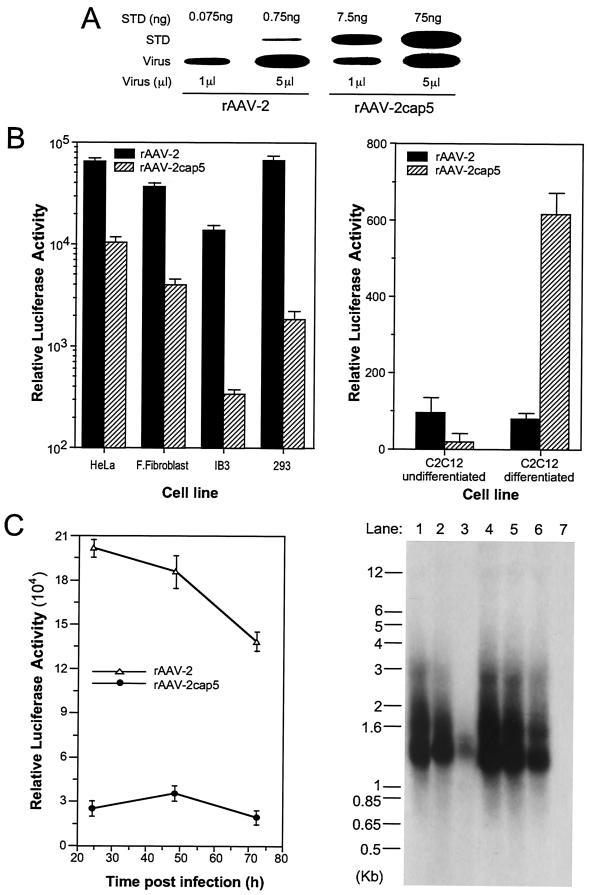
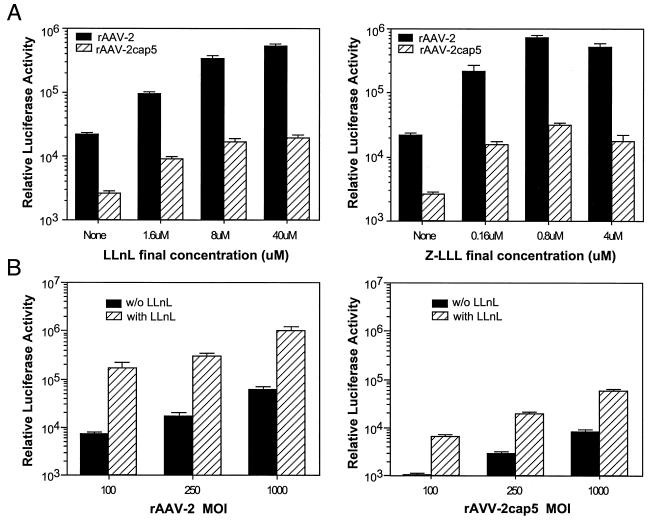
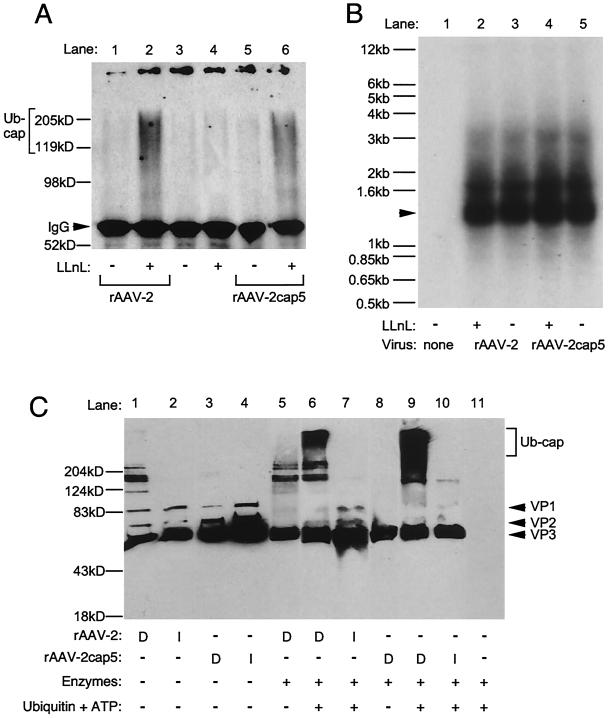
In the presence of complementing adeno-associated virus type 2 (AAV-2) Rep proteins, AAV-2 genomes can be pseudotyped with the AAV-5 capsid to assemble infectious virions. Using this pseudotyping strategy, the involvement of the ubiquitin-proteasome system in AAV-5 and AAV-2 capsid-mediated infections was compared. A recombinant AAV-2 (rAAV-2) proviral luciferase construct was packaged into both AAV-2 and AAV-5 capsid particles, and transduction efficiencies in a number of cell lines were compared. Using luciferase expression as the end point, we demonstrated that coadministration of the viruses with proteasome inhibitors not only increased the transduction efficiency of rAAV-2, as previously reported, but also augmented rAAV-5-mediated gene transfer. Increased transgene expression was independent of viral genome stability, since there was no significant difference in the amounts of internalized viral DNA in the presence or absence of proteasome inhibitors. Western blot assays of immunoprecipitated viral capsid proteins from infected HeLa cell lysates and in vitro reconstitution experiments revealed evidence for ubiquitin conjugation of both AAV-2 and AAV-5 capsids. Interestingly, heat-denatured virus particles were preferential substrates for in vitro ubiquitination, suggesting that endosomal processing of the viral capsid proteins is a prelude to ubiquitination. Furthermore, ubiquitination may be a signal for processing of the capsid at the time of virion disassembly. These studies suggest that the previously reported influences of the ubiquitin-proteasome system on rAAV-2 transduction are also active for rAAV-5 and provide a clearer mechanistic framework for understanding the functional significance of ubiquitination.
Figures
Similar articles
-
Endosomal processing limits gene transfer to polarized airway epithelia by adeno-associated virus.J Clin Invest. 2000 Jun;105(11):1573-87. doi: 10.1172/JCI8317. J Clin Invest. 2000. PMID: 10841516 Free PMC article.
-
Cross-dressing the virion: the transcapsidation of adeno-associated virus serotypes functionally defines subgroups.J Virol. 2004 May;78(9):4421-32. doi: 10.1128/jvi.78.9.4421-4432.2004. J Virol. 2004. PMID: 15078923 Free PMC article.
-
Intracellular trafficking of adeno-associated virus vectors: routing to the late endosomal compartment and proteasome degradation.J Virol. 2001 Feb;75(4):1824-33. doi: 10.1128/JVI.75.4.1824-1833.2001. J Virol. 2001. PMID: 11160681 Free PMC article.
-
Evaluation of risks related to the use of adeno-associated virus-based vectors.Curr Gene Ther. 2003 Dec;3(6):545-65. doi: 10.2174/1566523034578131. Curr Gene Ther. 2003. PMID: 14683451 Review.
-
[Ubiquitination of recombinant adeno-associated viral vector and its application].Zhejiang Da Xue Xue Bao Yi Xue Ban. 2012 Sep;41(5):586-91. doi: 10.3785/j.issn.1008-9292.2012.05.020. Zhejiang Da Xue Xue Bao Yi Xue Ban. 2012. PMID: 23086655 Review. Chinese.
Cited by
-
Effects of Altering HSPG Binding and Capsid Hydrophilicity on Retinal Transduction by AAV.J Virol. 2021 Apr 26;95(10):e02440-20. doi: 10.1128/JVI.02440-20. Epub 2021 Mar 3. J Virol. 2021. PMID: 33658343 Free PMC article.
-
Inducible caspase 9-mediated suicide gene therapy using AAV6 vectors in a murine model of breast cancer.Mol Ther Methods Clin Dev. 2023 Nov 24;31:101166. doi: 10.1016/j.omtm.2023.101166. eCollection 2023 Dec 14. Mol Ther Methods Clin Dev. 2023. PMID: 38149057 Free PMC article.
-
Direct and retrograde transduction of nigral neurons with AAV6, 8, and 9 and intraneuronal persistence of viral particles.Hum Gene Ther. 2013 Jun;24(6):613-29. doi: 10.1089/hum.2012.174. Hum Gene Ther. 2013. PMID: 23600720 Free PMC article.
-
Distinct transduction difference between adeno-associated virus type 1 and type 6 vectors in human polarized airway epithelia.Gene Ther. 2013 Mar;20(3):328-37. doi: 10.1038/gt.2012.46. Epub 2012 Jun 14. Gene Ther. 2013. PMID: 22695783 Free PMC article.
-
Scalable generation of high-titer recombinant adeno-associated virus type 5 in insect cells.J Virol. 2006 Feb;80(4):1874-85. doi: 10.1128/JVI.80.4.1874-1885.2006. J Virol. 2006. PMID: 16439543 Free PMC article.
References
-
- Chao, H., Y. Liu, J. Rabinowitz, C. Li, R. J. Samulski, and C. E. Walsh. 2000. Several log increase in therapeutic transgene delivery by distinct adeno-associated viral serotype vectors. Mol. Ther. 2:619-623. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
