

A replication-competent, neutralization-sensitive variant of simian immunodeficiency virus lacking 100 amino acids of envelope
- PMID: 11836385
- PMCID: PMC135934
- DOI: 10.1128/jvi.76.5.2075-2086.2002
A replication-competent, neutralization-sensitive variant of simian immunodeficiency virus lacking 100 amino acids of envelope
Abstract
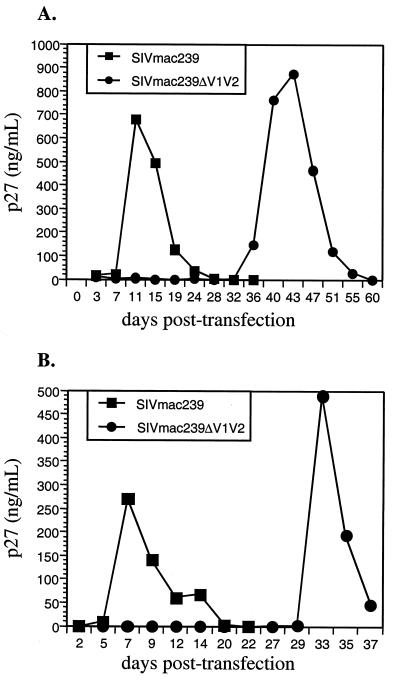
Coding sequences for the first two variable loops of the gp120 envelope glycoprotein were removed from simian immunodeficiency virus (SIV) strain 239 (SIVmac239). This deletion encompassed 100 amino acids. The resulting virus replicated poorly after transfection into immortalized T-cell lines, with peak replication occurring only after 25 to 30 days. Limited passaging of SIVmac239DeltaV1V2 in cultures gave rise to a variant which had significantly improved replication kinetics but which retained the original 100-amino-acid deletion in gp120. Cloning and sequencing revealed 11 changes in the envelope, including amino acid substitutions in both gp120 (5 substitutions) and gp41(6 substitutions). Four of the five changes in gp120 are predicted to lie within and around the putative coreceptor binding domain, a region which is believed to be covered by the V1 and V2 loops in the native envelope complex. Analysis of recombinant clones surprisingly revealed that the changes in gp41 were sufficient to overcome the replication deficiency created by deletion of the V1 and V2 loops from gp120. The SIVmac239DeltaV1V2 envelope displayed a significant reduction in its ability to mediate cell-cell fusion, and the infectious titer of SIVmac239DeltaV1V2 was approximately four- to eightfold lower than that of parental SIVmac239. Although SIVmac239 is strongly dependent on both CD4 and a coreceptor for entry, envelope protein lacking the V1 and V2 loops was able to mediate fusion with CD4(-) CCR5(+) cells at 60% the level observed with CD4(+) CCR5(+) cells. Plasma from SIVmac239-infected monkeys was at least 100 to 1,000 times more effective at neutralizing SIVmac239DeltaV1V2 than SIVmac239. These results demonstrate the dispensability of the V1-V2 sequences of SIVmac239 for viral replication, a role for V1 and V2 in shielding the coreceptor binding region of the envelope, and the extreme sensitivity of a SIV lacking these sequences to antibody-mediated neutralization.
Figures







References
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1987. Current protocols in molecular biology. John Wiley & Sons, Inc., New York, N.Y.
-
- Bandres, J. C., Q. F. Wang, J. O'Leary, F. Baleaux, A. Amara, J. A. Hoxie, S. Zolla-Pazner, and M. K. Gorny. 1998. Human immunodeficiency virus (HIV) envelope binds to CXCR4 independently of CD4, and binding can be enhanced by interaction with soluble CD4 or by HIV envelope deglycosylation. J. Virol. 72:2500-2504. - PMC - PubMed
-
- Bosch, M. L., P. L. Earl, K. Fargnoli, S. Picciafuoco, F. Giombini, F. Wong-Staal, and G. Franchini. 1989. Identification of the fusion peptide of primate immunodeficiency viruses. Science 244:694-697. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 35383/PHS HHS/United States
- RR 00168/RR/NCRR NIH HHS/United States
- P30 AI042845/AI/NIAID NIH HHS/United States
- R37 AI025328/AI/NIAID NIH HHS/United States
- P51 RR000168/RR/NCRR NIH HHS/United States
- K26 RR000168/RR/NCRR NIH HHS/United States
- R01 AI025328/AI/NIAID NIH HHS/United States
- AI 25328/AI/NIAID NIH HHS/United States
- F32 AI 10577/AI/NIAID NIH HHS/United States
- AI 42845/AI/NIAID NIH HHS/United States
- AI 35365/AI/NIAID NIH HHS/United States
- F32 AI010577/AI/NIAID NIH HHS/United States
- P01 AI035365/AI/NIAID NIH HHS/United States
- U01 AI035365/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
