

doi: 10.1128/jvi.76.5.2567-2572.2002.
Sequence variation in a newly identified HLA-B35-restricted epitope in the influenza A virus nucleoprotein associated with escape from cytotoxic T lymphocytes
Affiliations
- PMID: 11836437
- PMCID: PMC153818
- DOI: 10.1128/jvi.76.5.2567-2572.2002
Item in Clipboard
Sequence variation in a newly identified HLA-B35-restricted epitope in the influenza A virus nucleoprotein associated with escape from cytotoxic T lymphocytes
J Virol.
2002 Mar.
Abstract
Here, we describe a new HLA-B*3501-restricted cytotoxic T lymphocyte (CTL) epitope in the influenza A virus (H3N2) nucleoprotein, which was found to exhibit a high degree of variation at nonanchor residues. The influenza virus variants emerged in chronological order, and CTLs directed against old variants failed to recognize more recent strains of influenza A virus, indicating an escape from CTL immunity.
Figures
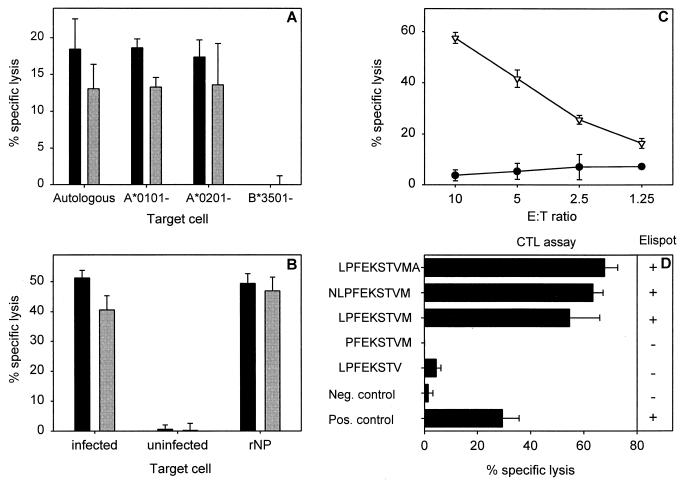
Characterization of an HLA-B*3501-restricted epitope and CTL clone (1980-1). (A) HLA class I restriction of CTL clone 1980-1 was determined by partially mismatched target cells (BLCL) infected with influenza A (H3N2) virus (Resvir-9) in a 51Cr release assay at effector-to-target cell (E:T) ratios of 10:1 (black bars) and 5:1 (grey bars) as described previously (3). Autologous BLCL cells (HLA-A*0101, HLA-A*0201, HLA-B*0801, and HLA-B*3501) and BLCL cells mismatched for a single HLA class I molecule (HLA-A*0101−, HLA-A*0201−, and HLA-B*3501−) were used. All BLCL cells expressed HLA-B*0801. The percent lysis of uninfected target cells was subtracted from the percent lysis of target cells infected with influenza virus. (B) Protein specificity of an HLA-B*3501-restricted CTL clone. HLA-B*3501+ target cells were infected with influenza A (H3N2) virus (Resvir-9) or were left uninfected and used as positive and negative controls, respectively. In addition, HLA-B*3501+ target cells were incubated overnight with 100 μg of bacterially expressed rNP of A/Netherlands/18/94 (A/H3N2)/ml as previously described (33). The target cells were used in a 51Cr release assay with CTL clone 1980-1 at E:T ratios of 10:1 (black bars) and 5:1 (grey bars). (C) A 9-mer peptide (NP418-426; LPFEKSTVM) (▿), predicted with an HLA binding prediction program (25), was loaded onto HLA-B*3501+ BLCL cells and used as target cells in a 51Cr release assay with the T-cell clone 1980-1 at the indicated E:T ratios. Untreated BLCL cells (•) were included as a negative control. (D) Minimal epitope mapping of HLA-B*3501-restricted epitope. Peptides were synthesized based on the initial 9-amino-acid sequence LPFEKSTVM that was extended or truncated at the C- or N-terminal end of the NP418-426 epitope in order to determine the minimal epitope. BLCL cells loaded with 5 μM concentrations of the different peptides were used as target cells in a CTL assay at an E:T ratio of 5:1 or applied to stimulate the CTL clone (1980-1) in an enzyme-linked immunospot assay. A plus indicates gamma interferon (IFN-γ) production by the CTL clone, while a minus indicates no production of IFN-γ. Influenza virus (Resvir-9)-infected BLCL cells and uninfected BLCL cells were included in both assays as positive and negative controls, respectively. Percent lysis is given as the mean ± standard deviation, and the results are representative of multiple experiments.
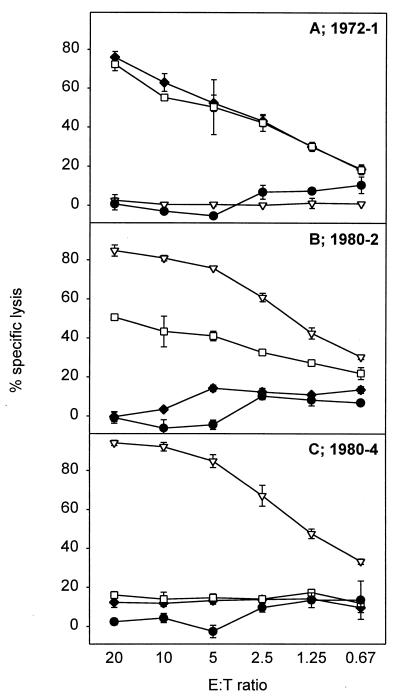
NP418-426 variant epitope specificity of CTL clones. Target cells (BLCL) were incubated overnight with 5 μM concentrations of the HLA-B*3501-restricted NP418-426 epitope variants, namely, NP418-426/1980 (▿), NP418-426/1972 (□), and NP418-426/1957 (⧫), or were left untreated (•) and used as a negative control. Clones 1972-1 (A), 1980-2 (B), and 1980-4 (C), obtained from different donors, were added at different effector-to-target cell ratios, and specific lysis was calculated. The results, given as the percent specific lysis (mean ± standard deviation), are representative of multiple assays.
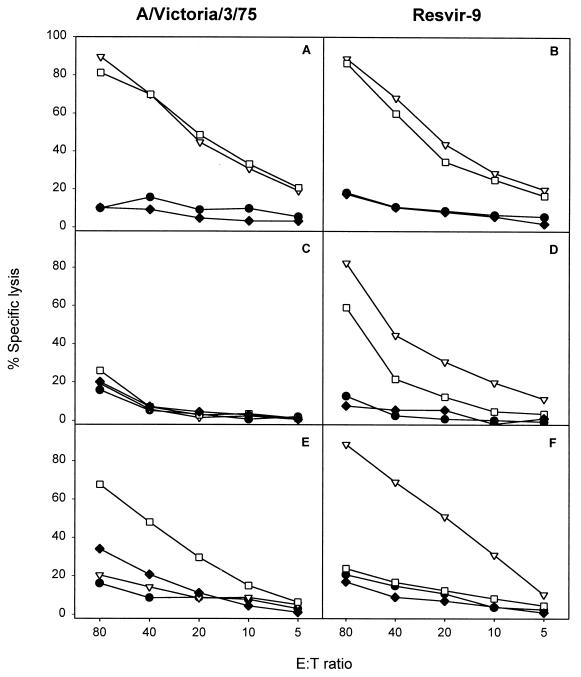
Epitope specificity of the polyclonal response following in vitro stimulation of PBMC cryopreserved in the year 2001 with influenza A viruses (H3N2) containing different variants of the epitope. PBMC of HLA-B35+ donors, aged between 30 and 50 years, were stimulated in vitro with A/Victoria/3/75 (A, C, and E), containing the NP418-426/1972 variant, or Resvir-9 (B, D, and F), containing the NP418-426/1980 epitope, as described previously (3). After 8 days, the effector cells were tested for lytic activity against target cells incubated with the peptides corresponding to NP418-426/1980 (▿), NP418-426/1972 (□), and NP418-426/1957 (⧫). Untreated target cells (•) were included in each assay as a negative control. Mean percentages of specific lyses of two independently repeated experiments are shown for donors 5972 (A and B), 5017 (C and D), and 5991 (E and F).
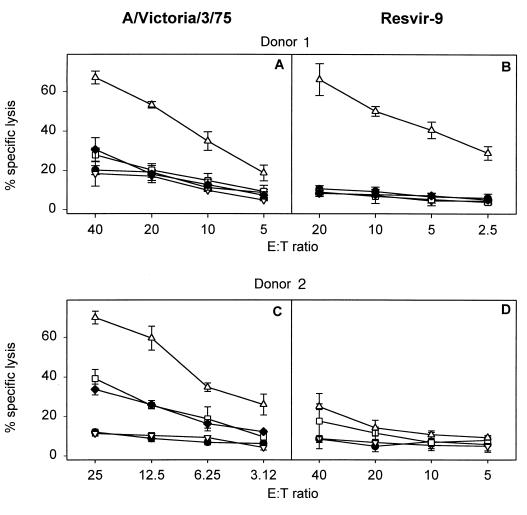
CTL responsiveness after stimulation of PBMC cryopreserved between 1982 and 1984 with influenza A viruses (H3N2) containing the NP418-426/1980 or NP418-426/1972 variant epitope. PBMC from two HLA-A2+ HLA-B35+ donors (donors 1 and 2) were stimulated in vitro with A/Victoria/3/75 (A and C), containing the NP418-426/1972 variant epitope, or Resvir-9 (B and D), containing the NP418-426/1980 variant epitope. The lytic activities of the effector cells were determined after 8 days of culture against target cells incubated with 5 μM concentrations of peptides corresponding to NP418-426/1980 (▿), NP418-426/1972 (□), NP418-426/1957 (⧫), or M158-66 epitope (▵), and a negative control (•) at different effector-to-target cell ratios. NP418-426/1957 was not determined for PBMC of donor 2 after stimulation with Resvir-9. The mean percentage of specific lysis ± standard deviation is given for one experiment.
References
-
- Bertoletti, A., A. Costanzo, F. V. Chisari, M. Levrero, M. Artini, A. Sette, A. Penna, T. Giuberti, F. Fiaccadori, and C. Ferrari. 1994. Cytotoxic T lymphocyte response to a wild type hepatitis B virus epitope in patients chronically infected by variant viruses carrying substitutions within the epitope. J. Exp. Med. 180:933-943. - PMC - PubMed
-
- Bertoletti, A., A. Sette, F. V. Chisari, A. Penna, M. Levrero, M. De Carli, F. Fiaccadori, and C. Ferrari. 1994. Natural variants of cytotoxic epitopes are T-cell receptor antagonists for antiviral cytotoxic T cells. Nature 369:407-410. - PubMed
-
- Boon, A. C. M., G. de Mutsert, Y. M. F. Graus, R. A. M. Fouchier, K. Sintnicolaas, A. D. M. E. Osterhaus, and G. F. Rimmelzwaan. 2002. The magnitude and specificity of influenza A virus-specific cytotoxic T-lymphocyte responses in humans is related to HLA-A and -B phenotype. J. Virol. 76:582-590. - PMC - PubMed
-
- Borrow, P., H. Lewicki, X. Wei, M. S. Horwitz, N. Peffer, H. Meyers, J. A. Nelson, J. E. Gairin, B. H. Hahn, M. B. Oldstone, and G. M. Shaw. 1997. Antiviral pressure exerted by HIV-1-specific cytotoxic T lymphocytes (CTLs) during primary infection demonstrated by rapid selection of CTL escape virus. Nat. Med. 3:205-211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
